
Проблемы Эволюции |
Совместный бакалавриат ВШЭ и РЭШ. Курс лекций.
предыдущий раздел следующий раздел оглавление
© А. В. Марков, 2011
Курс лекций
ДЗ. Прочесть «Происхождение видов» Дарвина. Отнестись с уважением, но критически. Написать эссе (1000 – 1500 слов) на тему «Взгляды Дарвина на ......................... с точки зрения современной науки». Вместо многоточия нужно вписать любую интересную и не слишком частную проблему, о которой идет речь в книге. Примеры таких проблем: наследственность, изменчивость, искусственный отбор, естественный отбор, борьба за существование, наследование результатов упражнения органов, половой отбор, происхождение сложных органов, соотношение постепенности и скачкообразности в эволюции, проблема переходных форм, проблема вида в биологии, развитие адаптаций, причины несовершенства организмов, биогеографические следствия эволюции и т.д. и т.п.). Это должен быть аналитический обзор, т.е. пересказ идей Дарвина (по тексту книги, так, чтобы было понятно, что вы ее читали) + критический анализ этих идей с точки зрения того, что нам известно сегодня. Можно опираться на любые заслуживающие доверия источники.
Как правило, мысли Дарвина относятся к одной из следующих категорий:
«Origin of species by means of Natural Selection and all sorts of other things»
Каждый ген в генофонде популяции может существовать в виде нескольких вариантов – аллелей. В результате мутаций возникают новые аллели (старых генов). Термины «мутация» и «аллель» генетики часто смешивают. Это жаргон, но все привыкли. Поэтому часто говорят «мутация распространилась в популяции», хотя более правильно – «аллель распространился в популяции».
Нейтральные мутации – не влияют на приспособленность.
Приспособленность – это эффективность размножения.
Например, если каждый носитель аллеля а1 оставляет в среднем 5 жизнеспособных потомков, а каждый носитель аллеля а2 (т.е. другого варианта того же гена А) оставляет в среднем 4 столь же жизнеспособных потомков, то приспособленность аллеля А1 по отношению к аллелю А2 = 5/4 = 1.25
Fa1/Fa2 = 1.25
Как правило, абсолютную приспособленность не измеряют – это не очень понятно, как сделать. А вот относительную приспособленность померить можно: просто посмотреть, с какой скоростью размножаются носители данного аллеля по сравнению со всеми остальными (т.е. с теми, у кого этого аллеля нет).
Различие между аллелями а1 и а2 – не нейтральное. Аллель а1 – «лучше», чем а2 (он повышает приспособленность).
Если же Fa1/Fa2 ≈ 1, то это нейтральное различие. Если в генофонде был сначала только аллель а1, а потом в результате мутации появился аллель а2, причем Fa1/Fa2 ≈ 1, то это значит, что мутация, в результате которой возник аллель а2, была нейтральной.
Разберемся, как будет меняться частота нейтральной мутации (нейтрального аллеля) со временем, с течением поколений.
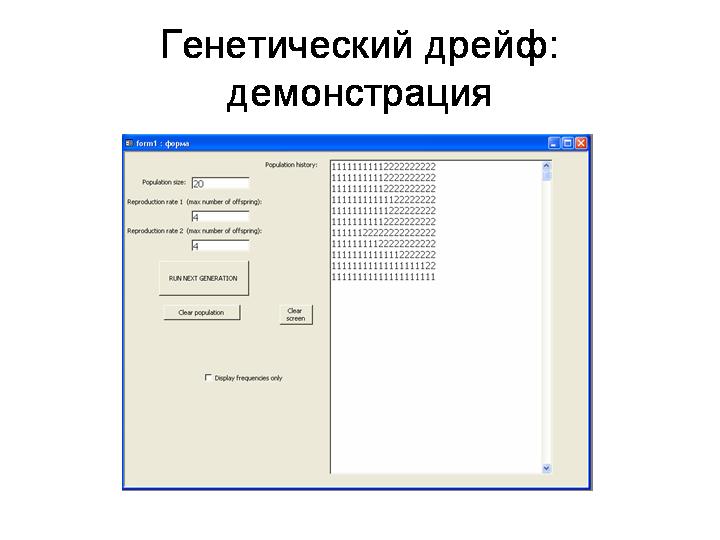
Демонстрация на модели «Genetic drift» (при параметрах 20, 4, 4). Устройство модели: Популяция из N гаплоидных особей. Два нейтральных аллеля: 1 и 2. В начальный момент времени частоты аллелей равны (q1=q2=0.5). Каждая особь производит потомков (случайное число в диапазоне от 0 до укузанного числа). Из этих потомков затем случайным образом отбирается N «выживших», которые и формируют следующее поколение в модельной популяции.
Выводы:
Формула 1. Чему равна вероятность фиксации (Pfix) нейтральной мутации? Если исходная частота q=0.5, то совершенно ясно, что аллель с равной вероятностью зафиксируется или элиминируется, т.е. Pfix = 0.5.
Представим себе ситуацию, когда у нас исходно есть не 2, а 4 нейтральных аллеля с одинаковыми частотами: q1=q2=q3=q4=0.25
Легко увидеть, что конечным результатом дрейфа будет фиксация одного из аллелей и элиминация трех других. Это становится ясно, если понять, что мы имеем полное право временно и условно объединить три из четырех аллелей в один, комплексный. Например, давайте считать, что у нас есть два аллеля: 1 и не-1. Аллель «не-1» – это общее название для аллелей 2, 3 и 4. Про два аллеля мы уже знаем, что один из них рано или поздно обязательно зафиксируется, а другой элиминируется. Стало быть, у нас либо зафиксируется аллель 1, либо «не-1». В первом случае все ясно – один аллель зафиксировался, три элиминировались. Во втором – у нас осталось три аллеля (2, 3 и 4). Давайте представим эту ситуацию в виде двух аллелей: 2 и не-2. И так далее.
Таким образом, можно считать доказанным, что конечным результатом дрейфа будет фиксация одного из аллелей и элиминация трех других. Ясно, что, поскольку исходно все 4 аллеля были в абсолютно равном положении, шансы на фиксацию у них должны быть одинаковы. В сумме эти шансы дают 1 (один из аллелей точно зафиксируется).
Следовательно, Pfix1 = Pfix2 = Pfix3 = Pfix4 = ј = 0.25
Вероятность фиксации нейтрального аллеля равна его исходной частоте (т.е. частоте в тот момент, для которого мы пытаемся рассчитать вероятность).
Поскольку мы могли с тем же успехом взять не 2, не 4, а любое другое число нейтральных аллелей, очевидно, что:
Pfix = q
Вероятность фиксации нейтрального аллеля равна его частоте. Это первая из трех формул, которую мы с вами должны вывести. А чему равна вероятность элиминации аллеля? Очевидно, 1-q.
Формула 2. Следующий очень важный вопрос – определить, сколько нейтральных мутаций будет фиксироваться в популяции в каждом поколении. Если мы сможем это установить, то мы обретем великий инструмент – молекулярные часы. Зная, с какой скоростью фиксируются в популяции нейтральные мутации, и зная, сколько их накопилось за какой-то период, мы сможем вычислить длительность этого периода. Например, сколько разных нейтральных мутаций есть в геномах человека и шимпанзе? Это можно подсчитать. Если бы мы еще знали, с какой скоростью фиксируются нейтральные мутации, мы могли бы на основе данных о числе накопившихся мутаций вычислить, как давно разошлись эволюционные линии предков человека и шимпанзе, понять, когда жил их последний общий предок.
Итак, сколько же нейтральных мутаций будет фиксироваться в популяции за одно поколение? Вывод этой формулы – это абсолютнейший шедевр биологической математики. Надеюсь, вы оцените.
Итак, от чего зависит скорость накопления нейтральных мутаций?
Очевидно, она зависит от того, с какой скоростью они появляются – то есть от скорости нейтрального мутагенеза, а также от того, какая часть вновь появляющихся нейтральных мутаций будет фиксироваться (а не элиминироваться).
Обозначим переменные:
Приступим к выводу формулы.
Очевидно, что V = M• Pfix
Искомая величина V равна числу мутаций, появляющихся за поколение, умноженному на ту долю, которая зафиксируется из них в конечном счете.
Чему равно M? Очевидно, что M = N•m
Это, надеюсь, ясно без пояснений.
А как найти Pfix, т.е. как выразить эту величину через те величины, которые можно измерить? Ну, мы ведь уже знаем, что Pfix = q , т.е. для каждой мутации вероятность зафиксироваться равна частоте этой мутации в данный момент. Новая, только что появившаяся мутация, очевидно, изначально есть только у одной особи. То есть ее частота q=1/N. Следовательно, и вероятность ее фиксации равна 1/N. Следовательно, из всех новых мутаций зафиксируется их часть, равная 1/N. Вот вам и искомое значение Pfix:
Pfix = q = 1/N
Итак, имеем:
V = M• Pfix M = N•m Pfix =1/N
Подставляем в первую формулу две другие, получаем:
V = N•m/N
Численность популяции великолепным образом сокращается! Итого:
V = m
Это и есть Вторая Великая Формула, и, наверное, самый блистательный образчик «биологической математики». Скорость фиксации нейтральных мутаций не зависит от численности популяции! Она равна m, т.е. просто-напросто скорости мутагенеза, скорости появления нейтральных мутаций в расчете на одну особь.
Поэтому у нас есть молекулярные часы. Мы их имеем благодаря генетическому дрейфу и благодаря тому, что многие мутации нейтральны. Принцип молекулярных часов:
D = 2V•t
где D – количество нейтральных различий между двумя видами, t – время, прошедшее с тех пор, когда жил последний общий предок этих двух видов. Двойка тут появляется потому, что нейтральные мутации накапливались обоими видами. Каждый из них накопил за время t количество нейтральных мутаций, равное D/2.
----------------------------------------------
Дарвин (рекомендации по эссе).

Главный вклад Дарвина в умственное развитие человечества. До Дарвина нам был известен только ОДИН механизм возникновения сложных объектов, предназначенных для какой-то цели или функции – разумный дизайн. Пейли и часы. Все подобные объекты трактовались как результат чьего-то сознательного проектирования. Или придумывались некие невнятные дополнительные сущности – жизненная сила, стремление к совершенству и т.п. (ламаркизм, номогенез). Что с научной точки зрения ничуть не лучше гипотезы о разумном дизайне. Дарвин открыл другой механизм – механизм самопроизвольного возникновения сложных объектов, приспособленных для выполнения функций – объектов, которые выглядят (для нас) так, как будто бы их кто-то нарочно спроектировал.
Философские аспекты: Dennett “Darwin’s Dangerous Idea”. Физики и химики подтянулись позже. Вдохновленный успехом нелинейной неравновесной термодинамики, Эйген в нобелевской речи даже назвал Дарвина с его идеей естественного отбора «великим физиком». Лаплас. До Дарвина биологи нуждались «в этой гипотезе». После Дарвина они могут без нее обойтись.
Напоминание формул, выведенных на прошлом занятии.


Если в генофонде популяции есть два аллеля с разным влиянием на приспособленность, то «более приспособленный» аллель в тенденции будет вытеснять «менее приспособленный» аллель из генофонда. Этот процесс называют аллельным замещением. По сути дела это конкурентное вытеснение одним аллелем другого. Это и есть естественный отбор в своем самом первичном, базовом виде, «очищенный от примесей».
Аллели (варианты) одного и того же гена конкурируют за доминирование в генофонде. Они как бы борются за место в хромосоме. Конечно, это метафорический язык. На самом деле, конечно, нет никакой сознательной конкуренции. Просто автоматически, в силу разного влияния аллелей на приспособленность организма, одних аллелей постепенно становится больше, а других – меньше. Но выглядит все так, как будто они конкурируют. Нам удобно так думать и удобно так говорить.
Кстати, и в экономике, мне кажется, конкуренция вовсе не обязана быть сознательной. Более эффективный процесс будет автоматически вытеснять менее эффективный процесс, если обоим требуются одни и те же ресурсы.
Два базовых эволюционных процесса: генетический дрейф (с которым мы уже познакомились) и аллельное замещение. Случайные и направленные изменения частот аллелей.
Как будут вести себя частоты аллелей в этом случае?
ДЕМОНСТРАЦИЯ НА МОДЕЛИ с параметрами 500, 20, 21 (5-процентная разница в приспособленности).
При численности 500 пятипроцентная разница в приспособленности обеспечивает фиксацию «хорошего» аллеля, хотя рост его частоты не является абсолютно неуклонным, в нем сохраняются элементы случайных блужданий.
Если мы увеличим N до 2000, рост q становится гораздо более гладким. Элемент случайности уменьшается (ДЕМОНСТРАЦИЯ). При N=4000 уже почти совсем неуклонный, гладкий рост частоты «хорошего» аллеля.
А что будет происходить в маленькой популяции? При N=20 «хороший» аллель начинает вести себя фактически как нейтральный! Отбор в такой маленькой популяции «не чувствует» пятипроцентной разницы в приспособленности (ДЕМОНСТРАЦИЯ).
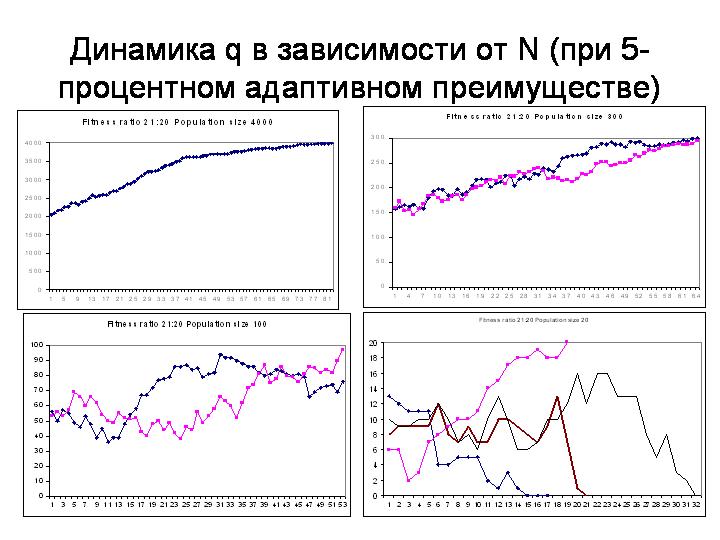
Здесь показано поведение q при одинаковом, пятипроцентном преимуществе в приспособленности, в разных по размеру популяциях.
Вывод: Численность популяции влияет на эффективность отбора. В маленькой популяции слабополезные и слабовредные аллели начинают вести себя фактически как нейтральные. В результате плохие аллели могут (иногда) фиксироваться, а хорошие – элиминироваться. Отбор в маленькой популяции «не чувствует» небольших различий в приспособленности.
Чем больше N (размер популяции), тем чувствительнее отбор к небольшим различиям F, и тем меньше шансов у слабовредной мутации – зафиксироваться, а у слабополезной – элиминироваться.
Можно сказать и так: чем больше популяция, тем слабее в ней эффекты генетического дрейфа и тем сильнее эффекты отбора.
Вопрос на плюсик: в связи с вышеизложенным, почему крупные животные вымирают чаще, чем мелкие?

Забудем временно про дрейф и попробуем рассмотреть эффект отбора в чистом виде, без дрейфа. Для этого нужно представить себе очень большую популяцию.
Попробуем вывести формулу аллельного замещения для очень большой популяции. Как будет вести себя q (частота аллеля 1) при заданной величине Fотн = F1/F2 (отношение приспособленности аллеля 1 к приспособленности конкурирующего аллеля 2)?
Пусть популяция очень большая, так что эффектом дрейфа можно пренебречь. Пусть начальное значение q0=0.5.
Носители аллелей 1 и 2 рожают потомков. При этом на каждого потомка с алелем 2 будет приходится Fотн потомков с аллелем 1. Например, если Fотн =2, то есть носители аллеля 1 производят вдвое больше потомков, то, соответственно, в первом поколении будет вдвое больше потомков с аллелем 1, чем с аллелем 2 (поскольку изначально их было поровну). Такое же соотношение сохраняется и у взрослых особей первого поколения (потому что взрослые – это случайная выборка из молоди).
Стало быть, при Fотн =2 и q0=0.5 в нулевом поколении соотношение частот аллелей 1 и 2 было 1:1. В первом поколении оно стало 2:1. В четвертом, очевидно, оно станет 4:1, потом 8:1 и т.д.
Стало быть, для поколения X получаем вот что:
q/(1 – q ) = Fотнx •q0/(1 – q0)
Относительная частота аллеля в поколении номер X равна его начальной относительной частоте, умноженной на его относительную приспособленность в степени X.
Обозначим для простоты величину q0/(1 – q0) какой-то одной буквой, например, k. Выразим теперь собственно величину q:
q = (1 – q ) •Fотнx •k
q = Fотнx •k – q •Fотнx •k
q + q •Fотнx •k = Fотнx •k
q • (1 + Fотнx •k) = Fотнx •k

Таким образом, мы получили общую формулу для процесса аллельного замещения в большой популяции (в которой можно пренебречь дрейфом). Аллельное замещение – это основной, первичный, базовый эволюционный процесс наряду с дрейфом генов.
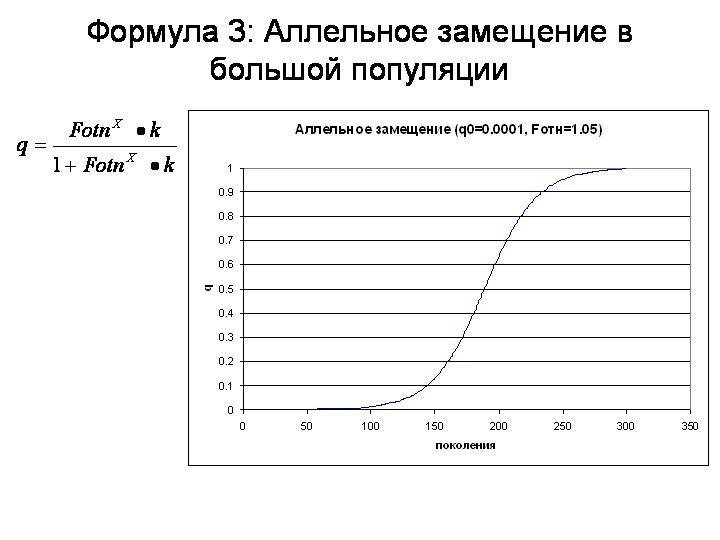
Вот как выглядит аллельное замещение на графике.
Очевидно, все это приложимо не только к биологии, к конкуренции между аллелями, но и к другим видам конкуренции. Я не знаю экономики, но подозреваю, что в общем случае процесс вытеснения чего-то менее эффективного чем-то более эффективным должен идти примерно так.
зачем нужен секс?
Почему и как появилось половое размножение – это одна из самых сложных тем в эволюционной биологии. Сотни идей и гипотез, аргументов и контраргументов. Основная проблема: затраты налицо, поэтому должен быть какой-то очень серьезный и быстрый выигрыш, иначе отбор не поддержит.
Гаплоидный и диплоидный жизненный цикл.
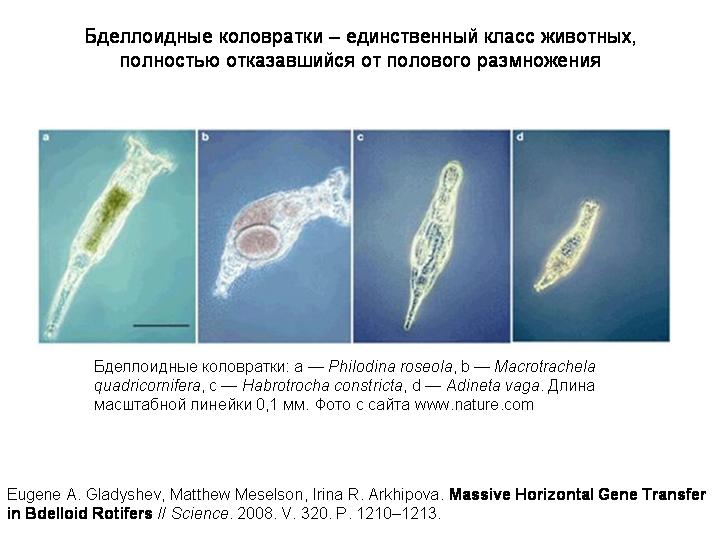
Начнем мы с того, что свяжем между собой половое размножение и другое явление, с которым мы уже знакомы – горизонтальный обмен генами. Как вы помните, у многоклеточных случаев горизонтального обмена генами известно мало. Однако в 2008 году было сделано потрясающее открытие, которое показало, что некоторые животные все-таки могут в массовом порядке заимствовать гены, причем не только у вирусов или МГЭ, а у клеточных организмов: у бактерий, грибов, растений.
Эти животные – бделлоидные коловратки, группа во многом уникальная (...)
Самая удивительная особенность бделлоидных коловраток – полный отказ от полового размножения. Я уже говорил о возможных параллелях между горизонтальным генетическим обменом и половым размножением, и о том, что половое размножение в определенном смысле заменило эукариотам горизонтальный генетический обмен. И вот на примере бделлоидных коловраток мы видим подверждение этой идеи.
Животные вообще довольно легко утрачивают половое размножение и переходят к партеногенезу (...). Такое не раз происходило в разных эволюционных линиях. Однако виды животных, отказавшиеся от полового размножения, имеют тенденцию очень быстро вымирать. Они не успевают дивергировать (...) и дать начало, допустим, целому бесполому семейству или отряду. Современные бесполые виды, как правило, представляют собой концевые веточки на эволюционном дереве. Это значит, что те бесполые виды, которые появлялись раньше, как правило, быстро вымирали и не успевали дать начало другим бесполым видам.
Бделлоидные коловратки представляют собой удивительное исключение. Это целый класс животных (включающий около 400 видов), которые размножаются только бесполым путем (партеногенетически). Никто никогда не видел самцов бделлоидных коловраток. По-видимому, полового размножения не было уже у общего предка бделлоидных коловраток, который жил много десятков миллионов, возможно, около 100 миллионов лет назад.
Бделлоидные коловратки — главный камень преткновения для всех теоретиков, пытающихся объяснить биологический смысл полового размножения. Какое ни придумай объяснение, сразу же возникает «проклятый» вопрос: если половое размножение такое полезное, как же бделлоидные коловратки без него обходятся? И если бделлоидные коловратки научились без него обходиться, почему другие животные не пошли по тому же пути?
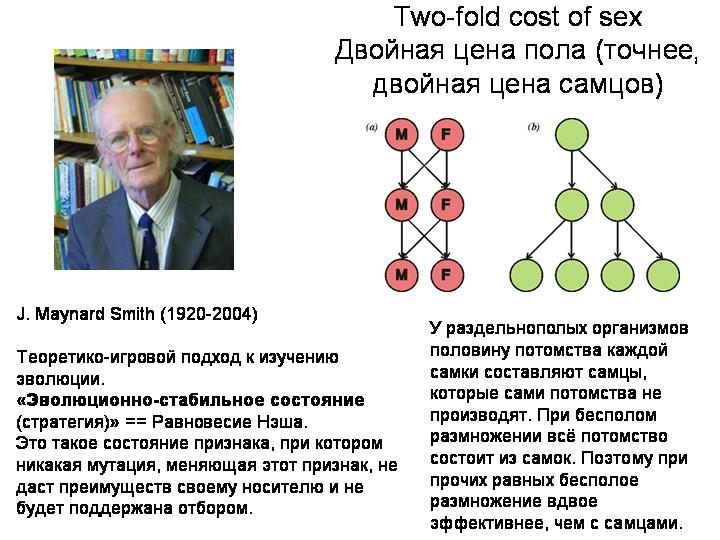
Ведь половое размножение — весьма «дорогое удовольствие» с точки зрения естественного отбора. При бесполом размножении вы передаете каждому потомку все свои гены, а при половом — только половину. За половое размножение приходится платить двукратным снижением эффективности передачи генов потомству.
Джон Мэйнард Смит назвал этот парадокс «двойной ценой пола». Здесь надо сделать небольшое отступление. Дж. Мэйнард Смит – один из величайших биологов-теоретиков 20 века. Одной из его заслуг является то, что он, вместе с некоторыми другими авторами, ввел в биологический обиход теорию игр и ввел такое понятие, как «эволюционно стабильное состояние» (или стратегия).
Эволюционно-стабильное состояние в биологии соответствует «равновесию Нэша» в теории игр. Равновесие Нэша — так в теории игр называется тип решений игры двух и более игроков, в котором ни один участник не может увеличить выигрыш, изменив своё решение (свою стратегию игры) в одностороннем порядке.
Соответственно, эволюционно-стабильное состояние – это такое состояние признака в популяции, при котором никакая мутация, меняющая этот признак, не даст преимуществ своему носителю и не будет поддержана отбором.
Много всего хорошего могло бы появиться в эволюции: например, всеобщая любовь и дружба между особями в каждой популяции, самопожертвование для общего блага и т.п. Но в большинстве случаев такие стратегии не являются эволюционно стабильными. Допустим, у нас есть вид животных, все особи которого друг за друга горой и готовы жертвовать жизнью ради спасения любых других представителей своего вида. Это было бы очень хорошо для вида в целом. Но эволюции нет дела до интересов вида. Такая ситуация не будет эволюционно стабильной, и вот почему.
Предположим, у нас есть такой вид, состоящий сплошь из героев-альтруистов. Что произойдет, если в ней появится мутант-эгоист? Он, к сожалению, почти наверняка получит адаптивное преимущество. Он будет пользоваться добротой других особей, а сам ничего не будет давать взамен. В общем случае его стратегия будет более выигрышной – он оставит больше потомства. Соответственно «ген эгоистичного поведения» будет распространяться! В итоге ген эгоизма может полностью вытеснить ген альтруизма. Иногда, при особых условиях, этого не происходит, но все-таки чаще, как правило, в природе живые существа ведут себя эгоистично, хоть это и плохо для вида. Это плохо для вида, но выгодно для самих эгоистичных особей и для их генов, которые благодаря этому эгоизму распространяются лучше, чем гены альтруистов.
У нас будет специальная тема «эволюция альтруизма», и мы подробно разберем ситуации, при которых альтруизм все-таки развивается. Но в общем случае, как правило, если нет каких-то особых условий, альтруизм – эволюционно нестабильная стратегия. Она вытесняется мутантами-эгоистами.
Вернемся к половому размножению. Половое размножение — «дорогое удовольствие» с точки зрения естественного отбора. Джон Мейнард Смит назвал центральную проблему, с которой сталкиваются двуполые виды – имеющие самцов и самок – проблемой «двойной цены пола». Суть идеи в том, что разделение на самцов и самок должно быть эволюционно нестабильным.
У раздельнополых организмов половину потомства каждой самки составляют самцы, которые сами потомства не производят. При бесполом размножении всё потомство состоит из самок. Поэтому при прочих равных бесполое размножение вдвое эффективнее, чем с самцами.
Следовательно, раздельнополость должна быть эволюционно нестабильна. Это не равновесие по Нэшу. Представьте, что у нас есть популяция двуполых животных, и в ней вдруг появляется мутантная самка, способная размножаться партеногенезом.
Потомство этой самки, очевидно, должно очень быстро вытеснить потомство всех остальных самок. Просто потому, что партеногенетические самки будут размножаться вдвое быстрее, чем самки, вынужденные спариваться с самцами и рожать не только дочерей, но и сыновей.
Предполагается, что каждая самка может произвести на свет ограниченное количество потомков – будь то сыновья или дочери (например, только двух, как на рисунке). Если она производит только дочерей, способных обходиться без самцов, ее потомство будет размножаться ровно вдвое быстрее.
В результате гены раздельнополых особей будут вытеснены генами партеногенетических особей. Это и значит, что раздельнополость эволюционно нестабильна.
Из этого, далее, следует, что если, несмотря на двойную цену самцов у многих организмов все-таки есть самцы, то это должно давать какое-то преимущество. Настолько значительное, что оно перевешивает эту двойную цену.
----------------------------------------------------
Теперь вернемся к бделлоидным коловраткам. Они отказались от полового размножения, что вроде бы соответствует идеям Мэйнарда Смита. Отказались, потому что это позволяет им не платить «двойную цену». Но, с другой стороны, они при этом должны были потерять и то преимущество, в чем бы оно ни заключалось, которое у других животных перевешивает двойную цену самцов. Многие авторы отмечали, что любое теоретическое построение, указывающее на преимущества полового размножения, обязательно должно как-то объяснять парадокс бделлоидных коловраток.
И вот, похоже, удалось нащупать решение этой давней загадки. В хромосомах бделлоидной коловратки обнаружено множество генов, не встречающихся ни у каких других животных. Некоторые из этих генов явно имеют бактериальное происхождение. Другие происходят от грибов, третьи — от растений. Были выявлены десятки генов, заимствованных коловратками у представителей других царств.
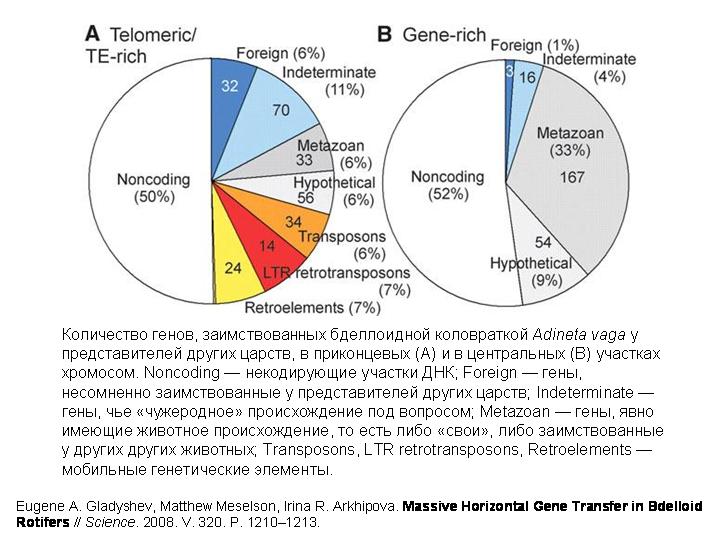
Некоторые заимствованные бактериальные гены у коловраток сохранили структуру, характерную для прокариотических генов (в них нет интронов), а другие уже успели обзавестись интронами, характерными для эукариот (как – неизвестно). По крайней мере некоторые из заимствованных генов реально работают в клетках коловраток и кодируют функциональные белки.
Примененные методы не позволяли выявить гены, заимствованные у других животных или, тем более, у других бделлоидных коловраток. Выявлялись только гены, заимствованные не у животных.
Животные стараются оберегать свои половые клетки от чужого генетического материала, в том числе вирусного. У бделлоидных коловраток эти барьеры, по-видимому, ослаблены. Это может быть связано с необычным образом жизни этих животных. Они живут в мелких лужах и отлично переносят высыхание. Потом их, как пыль, может перенести ветром в другую лужу. Однако при высыхании мембраны клеток могут повреждаться, что облегчает проникновение чужой ДНК. При высыхании образуются разрывы в хромосомах, которые клеткам приходится зашивать, репарировать, когда коловратка снова размокнет. В ходе репарации разорванных хромосом имеется вероятность случайного включения в хромосому чужеродного фрагмента.
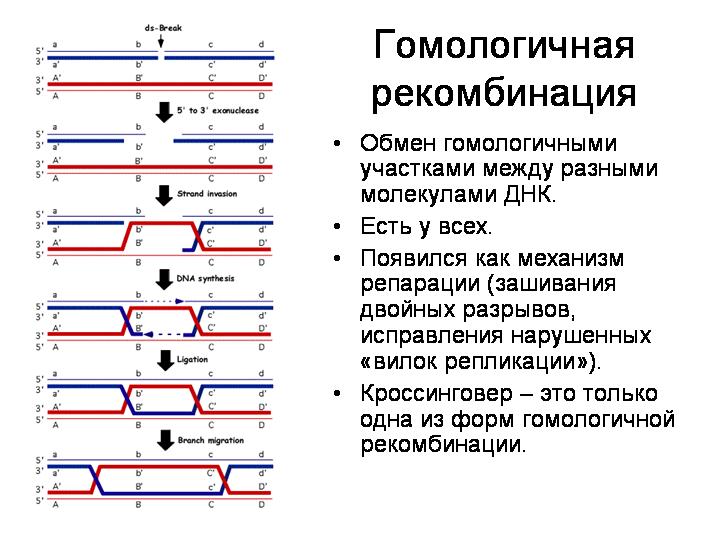
При высыхании образуются разрывы в ДНК, которые клеткам приходится репарировать при помощи гомологичной рекомбинации. При этом есть вероятность включения в хромосому чужеродного фрагмента.
Пока еще не известно, обмениваются ли бделлоидные коловратки генами между собой. Однако вряд ли коловратки, охотно заимствующие гены у микробов и растений, имеют при этом какую-то особую систему защиты от заимствования генов у родственников. Если же они хотя бы иногда меняются генами друг с другом, то получается, что они на самом деле не отказались от межорганизменной генетической рекомбинации, то есть перемешивания генов разных родителей в геноме потомства. Они просто вернулись от продвинутого варианта такой рекомбинации (полового процесса) к более примитивному варианту — горизонтальному обмену.

Этот пример подтверждает, что половой процесс и горизонтальный обмен генами – явления родственные. Судите сами: бделлоидные коловратки – единственная группа животных, которая давно отказалась от полового размножения и до сих пор не вымерла. С другой стороны, единственная группа животных, у которой обнаружен массовый горизонтальный обмен генами – это опять-таки бделлоидные коловратки. Такое совпадение не может быть случайным.
В чем же состоит это загадочное преимущество, которое дает половой процесс? Преимущество, утратив которое, виды, как правило, быстро вымирают. Преимущество, из-за которого большинство животных все-таки не уподобились бделлоидным коловраткам. Которое должно проявляться быстро, а не через миллион лет.

Рассмотрим две главные гипотезы. Первая известна под названием «храповик Мёллера». Храповик — это устройство, в котором ось может крутиться только в одну сторону. Суть в том, что если у бесполого организма возникает вредная мутация, его потомки уже не могут от нее избавиться. Она будет, как родовое проклятие, передаваться всем его потомкам вечно (если только не произойдет обратная мутация, а вероятность этого очень мала).
У бесполых организмов отбор может отбраковывать только целые геномы, но не отдельные гены. Если в бесполой популяции случайно погибает особь с наименьшим числом вредных мутаций, это означает необратимое ухудшение, необратимую потерю. При отсутствии рекомбинации (обмена гомологичными генами между особями) невозможно получить из сильно отягощенных мутациями геномов менее отягощенный.
Поэтому в череде поколений бесполых организмов может (при соблюдении определенных условий) происходить неуклонное накопление вредных мутаций. Одним из таких условий является большой размер генома. Чем больше в геноме генов, тем больше новых мутаций будет в среднем у каждого организма. Поэтому бесполые организмы не могут позволить себе иметь большие геномы – точнее, большие осмысленные части геномов.
Бесполые организмы не могут себе позволить иметь маленькие популяции (и крупные размеры), малое число потомков (т.к. только очень эффективный очищающий отбор может их спасти).
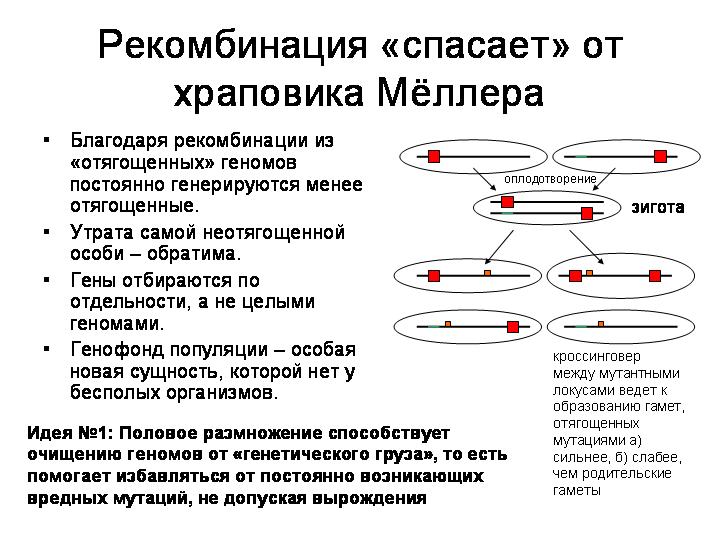
Если же организмы размножаются половым путем, то индивидуальные геномы постоянно рассыпаются и перемешиваются (в результате мейозов и слияний гамет), а новые геномы формируются из фрагментов, ранее принадлежавших разным организмам. В результате возникает особая новая сущность, которой нет у бесполых организмов, — генофонд популяции. Гены получают возможность размножаться или выбраковываться независимо друг от друга. Ген с неудачной мутацией может быть отбракован отбором, а остальные («хорошие») гены данного родительского организма могут благополучно сохраниться в популяции.
Таким образом, первая идея состоит в том, что половое размножение способствует очищению геномов от «генетического груза», то есть помогает избавляться от постоянно возникающих вредных мутаций, не допуская вырождения (снижения общей приспособленности популяции).
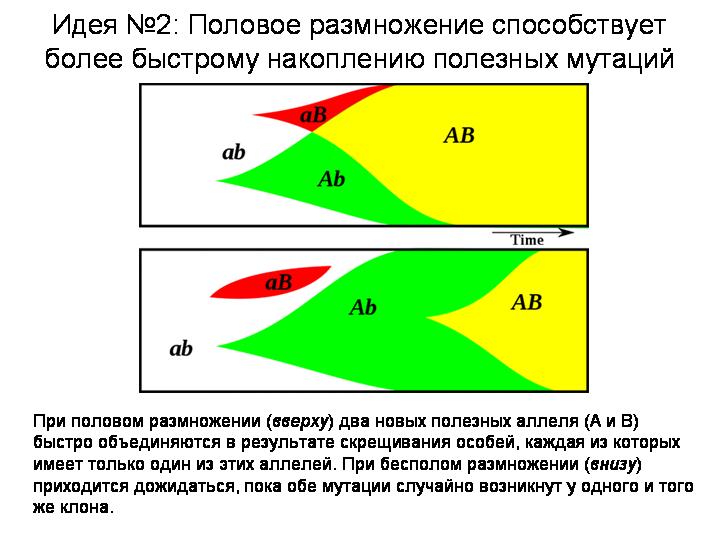
Вторая идея: половое размножение помогает эффективнее адаптироваться к меняющимся условиям за счет ускоренного накопления мутаций, полезных в данной обстановке. Допустим, у одной особи возникла одна полезная мутация, у другой — другая. Если эти организмы бесполые, у них мало шансов дождаться объединения обеих мутаций в одном геноме. Половое размножение дает такую возможность. Оно фактически делает все полезные мутации «общим достоянием». Ясно, что скорость приспособления к меняющимся условиям у организмов с половым размножением должна быть выше.
Все эти теоретические построения, однако, основаны на определенных допущениях. Результаты математического моделирования свидетельствуют о том, что степень полезности или вредности полового размножения по сравнению с бесполым размножением или самооплодотворением зависит от многих параметров. В их числе размер популяции; скорость мутирования; размер генома; количественное распределение мутаций в зависимости от степени их вредности/полезности; число потомков, производимых одной самкой; эффективность отбора (степень зависимости числа оставляемых потомков не от случайных, а от генетических факторов) и т. д. Некоторые из этих параметров очень трудно измерить не только в природных, но и в лабораторных популяциях.

Поэтому гипотезы остро нуждаются в экспериментальной проверке. Такие проверки были проведены. Рассмотрим одно исследование, выполненное на круглом черве Caenorhabditis elegans (биологи из университета штата Орегон, 2009 г). Это исследование показало действенность обоих механизмов, обеспечивающих преимущество тем популяциям, которые не отказываются от самцов, несмотря на их «двойную цену».
Уникальный объект для изучения роли самцов. Черви Caenorhabditis elegans словно нарочно созданы для проверки таких гипотез.
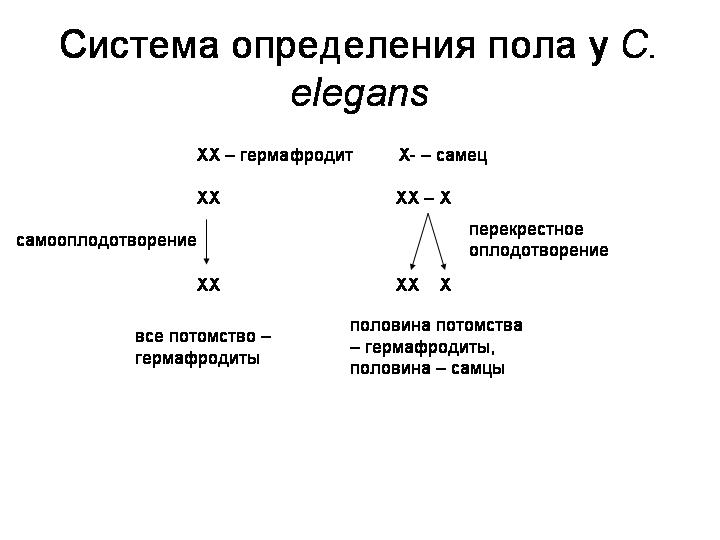
У этих червей нет самок. Популяции состоят из самцов и гермафродитов, причем последние преобладают. У гермафродитов две X-хромосомы, у самцов — только одна (система определения пола X0). Гермафродиты производят сперматозоиды и яйцеклетки и могут размножаться без посторонней помощи путем самооплодотворения. Самцы производят только сперматозоиды и могут оплодотворять гермафродитов. В результате самооплодотворения на свет появляются только гермафродиты. При перекрестном оплодотворении половина потомства оказывается гермафродитами, половина — самцами.
Обычно частота перекрестного оплодотворения в популяциях C. elegans не превышает нескольких процентов. Чтобы определить эту частоту, не обязательно наблюдать за интимной жизнью червей — достаточно знать процент самцов в популяции.
Следует пояснить, что самооплодотворение — это не совсем то же самое, что бесполое (клональное) размножение, однако различия между ними не очень существенны (они быстро сходят на нет в череде самооплодотворяющихся поколений. Самооплодотворяющиеся организмы за несколько поколений неизбежно становятся гомозиготными по всем локусам. После этого потомство перестает отличаться от родителей генетически, точно так же, как и при клональном размножении.
У C. elegans известны мутации, влияющие на частоту перекрестного оплодотворения. Одна из них смертельна для самцов и фактически приводит к тому, что в популяции остаются только гермафродиты, размножающиеся путем самооплодотворения. Другая мутация лишает гермафродитов способности производить сперму и фактически превращает их в самок. Популяция, в которой все особи несут эту мутацию, становится обычной раздельнополой популяцией, как у большинства животных.
Авторы сравнивали три породы червей с одинаковыми геномами, различающимися только наличием этих мутаций.
С этими породами провели два эксперимента.

В первом эксперименте проверялась гипотеза о том, что перекрестное оплодотворение помогает избавляться от «генетического груза». Эксперимент продолжался в течение 50 поколений (червей, а не экспериментаторов). Червей подвергали действию химического мутагена — этилметансульфоната. Это приводило к увеличению частоты мутирования в четыре раза. Молодых животных помещали в чашку Петри, разделенную пополам труднопреодолимой стенкой (см. на слайде), причем червей сажали в одну половину чашки, а их пища — бактерии E. coli — находилась в другой половине. Чтобы добраться до еды, а значит получить шанс выжить и оставить потомство, черви должны были преодолеть препятствие. Тем самым экспериментаторы повысили эффективность очищающего отбора, который отсеивает вредные мутации. В обычных лабораторных условиях эффективность отбора низкая, потому что черви окружены пищей со всех сторон. В такой ситуации могут выжить и размножиться даже очень слабые, перегруженные вредными мутациями животные. В новой экспериментальной установке этой уравниловке был положен конец. Чтобы переползти через стенку, червь должен быть здоров и крепок.
Авторы сравнивали приспособленность у червей до и после эксперимента, то есть у особей первого и пятидесятого поколения. Червей C. elegans можно хранить в замороженном виде. Это облегчает подобные эксперименты. Пока длился эксперимент, выборка червей 1-го поколения спокойно лежала в морозильнике. Приспособленность измеряли так. Червей смешивали в равной пропорции с контрольными червями, в геном которых был вставлен ген светящегося белка, и сажали в экспериментальную установку. Животным давали время, чтобы преодолеть барьер и размножиться, а затем определяли процент несветящихся особей в потомстве. Если этот процент увеличился в пятидесятом поколении по сравнению с первым — значит, в ходе эксперимента приспособленность выросла, если уменьшился — значит, имело место вырождение.
Результаты получились следующие. Подтвердилось предположение, что перекрестное оплодотворение помогает бороться с генетическим грузом. Чем выше частота перекрестного оплодотворения, тем лучше был итоговый результат (тем меньше выродились черви). Искусственно повышенная скорость мутирования привела к вырождению (снижению приспособленности) всех пород червей, кроме «облигатных перекрестников».
Самое интересное, что частота перекрестного оплодотворения у «диких» червей в ходе эксперимента стала заметно выше исходных 5%. Это означает, что в жестких условиях (имеется в виду как необходимость перелезать через барьер, так и повышенный темп мутагенеза) естественный отбор дает преимущество особям, размножающимся путем перекрестного оплодотворения. Потомство таких особей оказывается более жизнеспособным, и поэтому в ходе эксперимента происходит отбор на склонность к перекрестному оплодотворению.
Таким образом, первый эксперимент подтвердил гипотезу о том, что перекрестное оплодотворение помогает избавляться от вредных мутаций.

Во втором эксперименте проверялось, помогает ли перекрестное оплодотворение вырабатывать новые адаптации путем накопления полезных мутаций. На этот раз червям, чтобы добраться до пищи, нужно было преодолеть зону, заселенную патогенными бактериями Serratia. Эти бактерии, попадая в пищеварительный тракт C. elegans, вызывают у червя опасное заболевание. Чтобы выжить в этой ситуации, черви должны были либо научиться не глотать вредных бактерий, либо выработать устойчивость к ним. Какой из вариантов выбрали подопытные популяции — неизвестно, однако за 40 поколений черви облигатные перекрестники отлично приспособились к новым условиям, дикие черви приспособились хуже, а облигатные самооплодотворители не приспособились совсем (их выживаемость в среде с вредными бактериями осталась на исходном низком уровне). И снова в ходе эксперимента у диких червей под действием отбора возросла частота перекрестного оплодотворения.
Таким образом, перекрестное оплодотворение действительно помогает популяции приспосабливаться к новым условиям, в данном случае — к появлению болезнетворного микроба. Тот факт, что в ходе эксперимента у диких червей увеличивалась частота перекрестного оплодотворения, означает, что спаривание с самцами дает гермафродитам немедленное адаптивное преимущество, которое, очевидно, перевешивает «двойную цену», которую им приходится платить, производя на свет самцов.

Моделируется популяция гаплоидных организмов, размножающихся бесполым путем. 100, 3, 20, 0.05, 0.1, 0.7, 200
Параметры модели:
Population size – размер популяции (количество взрослых организмов в каждом поколении),
Reproduction rate – число потомков, производимых каждым взрослым организмом в каждом поколении (произведя потомков, взрослый организм погибает),
Number of housekeeping genes – Число «генов» (или генных комплексов), соответствующих каким-либо жизнеобеспечивающим функциям организма. У каждого гена каждого организма есть величина fitness (F, приспособленность), показывающая, с какой эффективностью данный ген выполняет свою функцию. Изначально все особи в популяции получают значения F=100 для каждого гена.
Mutation rate – частота мутирования (на 1 ген). Мутации возникают при образовании потомков. Потомок получает гены родительской особи, причем с вероятностью mutation rate каждый ген может мутировать. Мутация делятся на полезные и вредные. Полезные мутации увеличивают fitness гена на случайное число в диапазоне от 0 до 1, вредные – снижают fitness гена на случайное число в диапазоне от 0 до 5.
Beneficial mutation rate (BMR) – доля полезных мутаций (например, при Beneficial mutation rate = 0.1 каждая мутация с вероятностью 0.1 будет полезной, с вероятностью 0.9 – вредной). Таким образом, при заданных здесь параметрах «средний вред» мутаций превышает их «среднюю пользу» в 45 раз (вредные мутации возникают в 9 раз чаще и приносят в среднем в 5 раз больше вреда, чем полезные – пользы).
Selectivity of deaths (SD) – степень зависимости смертности потомства от генов, а не от случая. Например, при Selectivity of deaths = 0.7 гибель молодой особи на 70% определяется суммарной приспособленностью ее генов, на 30% - случайностью.
Все молодые особи, родившиеся в данном поколении, ранжируются в порядке убывания величины SD * (Fi - MinFit) / (MaxFit - MinFit) + Rnd * (1 - SD), где Fi – суммарная F всех геннов данной (i-ой) особи, MinFit – минимальная, а MaxFit – максимальная приспособленность молодых особей в данном поколении. Первые PS особей из данного списка становятся взрослыми, остальные погибают. Тем самым реализуется зависимость смертности от сочетания генетических и случайных причин, причем вклад генетических причин равен SD.
Предельный размер генома
Первое важное обстоятельство, которое демонстрируется этой моделью: Для каждого набора параметров существует предельный размер генома (NGmax), который не может быть увеличен (добавлением новых полезных генов) без неизбежного генетического вырождения (необратимого накопления вредных мутаций и снижения «приспособленности»). Я уже упоминал об этом, а модель это наглядно показывает.
Например, при этих значениях параметров NGmax ≈ 16.
ДЕМОНСТРАЦИЯ (показать, что при 15-16 генах все хорошо, а при 17-18 – уже плохо)
При наличии в геноме 17 или более «полезных генов» средняя приспособленность особей в популяции (Favg, средняя по всем генам и по всем особям) опускается ниже 100 и продолжает неуклонно снижаться с течением поколений. При NG ≤ 16 вырождения не происходит, средняя приспособленность не опускается ниже 100 (отбор справляется с отбраковкой вредных мутаций).
Это очень важный вывод. Существует максимальный размер полезной части генома, который не может быть превышен без неизбежного вырождения – при данном темпе мутирования. Это серьезнейшее ограничение для земной жизни.
Трансформация
Трансформация в модели реализуется так: с вероятностью TR каждый ген каждой взрослой особи заменяется случайно выбранным гомологичным геном любой другой взрослой особи в популяции. Предполагается, что микробы всасывают случайным образом ДНК из среды, причем в среде присутствуют гены, идентичные генам взрослых особей данного поколения, в соответствующей пропорции.
Уровень популяции
Модель показывает, что трансформация (замена одного или нескольких своих генов случайно выбранными гомологичными генами из генофонда популяции): 1) повышает эффективность положительного и отрицательного отбора, 2) позволяет организмам при той же скорости мутирования иметь больше генов в геноме, не подвергаясь генетическому вырождению.
Например, если при тех же параметрах позволить организмам осуществлять трансформацию, то наблюдается следующее:
ДЕМОНСТРАЦИЯ (20 генов; показать, что без трансформации – вырождение, от которого спасает даже очень небольшой (0.011) уровень трансформации). Но еще лучше – при замене 30-50% генов.
Показать, что с трансформацией 0,5 можно иметь 65 генов, а без нее – только 16.
Первые выводы:
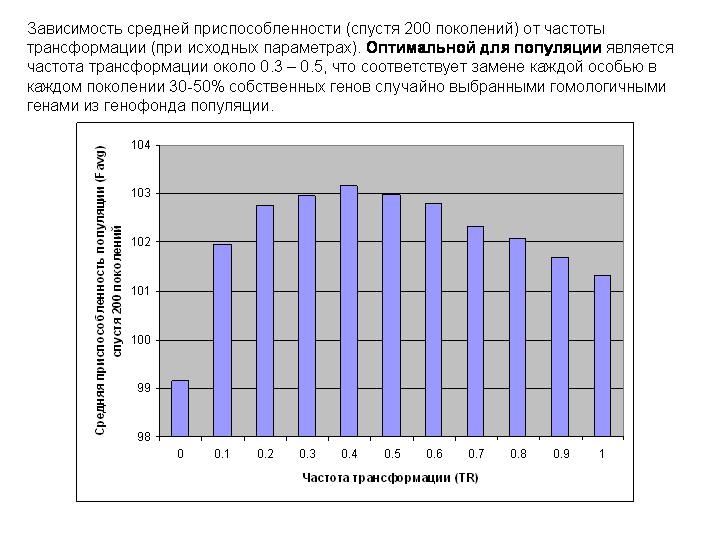
На слайде показана зависимость Favg (спустя 200 поколений) от TR (при тех же параметрах). Видно, что оптимальной для популяции является частота трансформации около 0.3 – 0.5, что соответствует замене каждой особью в каждом поколении 30-50% собственных генов случайно выбранными гомологичными генами других особей.
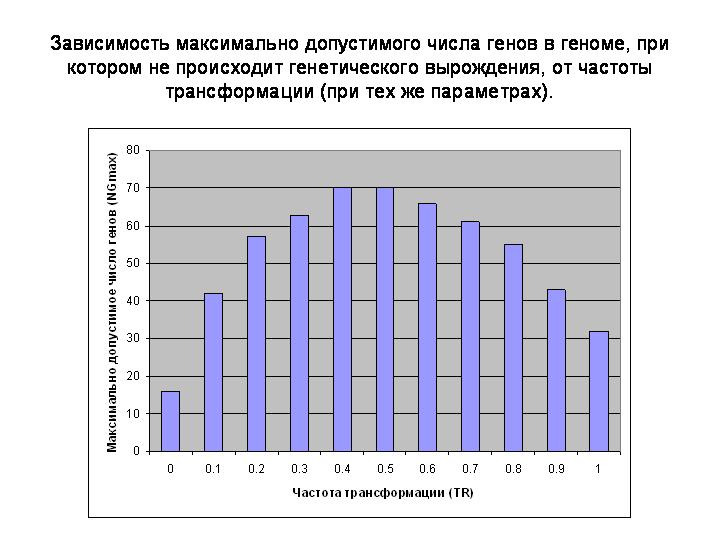
Здесь показана зависимость максимально допустимого числа генов в геноме, при котором не происходит генетического вырождения, от частоты трансформации.
Хорошо видно, что горизонтальный обмен позволяет организмам радикально увеличить число генов в геноме, не подвергаясь риску генетического вырождения. При данном наборе параметров без ГГО они могут позволить себе иметь лишь 16 генов, с небольшим ГГО (TR=0,1) это число возрастает до 40, а при TR = 0.4 - 0.5 до 70, т.е. более чем в 4 раза!
Таким образом, ГГО (трансформация) потенциально дает возможность:
Уровень особей (Индивидуальный, или организменный отбор).
Однако до сих пор мы рассматривали проблему с точки зрения «интересов вида» или «интересов популяции». Для популяции генетический обмен выгоден, это мы увидели. Но этого мало. Необходимо еще получить ответ на следующий вопрос: Зафиксируется ли «ген трансформации» в популяции микроорганизмов, или же микробы, способные к трансформации, будут систематически вытесняться из популяции микробами, к ней не способными? Такой вариант не исключен, потому что признак, полезный на уровне популяции, вполне может оказаться вредным на уровне особей и/или на уровне генов. И тогда этому признаку будет гораздо труднее появиться в результате эволюции. Иными словами, нужно вспомнить Мэйнарда Смита и проверить, будет ли трансформация эволюционно стабильным признаком.
ДЕМОНСТРАЦИЯ. Поместим в геном модельных организмов дополнительный ген, определяющий способность (или неспособность) осуществлять трансформацию. Этот ген не имеет своего значения F: полезность или вредность признака «способность к трансформации» будет выясняться в течение «жизни» модельной популяции, т.е. в ходе реализации модели.
Половине особей в исходной популяции был добавлен в геном ген tr+, обеспечивающий трансформацию с TR=0.5 (т.е. с оптимальной для популяции частотой). Другая половина особей получила нефункциональный аллель того же гена (tr-). Эти последние особи не могут осуществлять трансформацию.
Мы уже знаем, что для популяции было бы выгодно, чтобы особи tr+ вытеснили особей tr- Но «интересов популяции» недостаточно для того, чтобы особи tr+ реально смогли победить особей tr- в конкурентной борьбе внутри популяции.
В нашей модели при данном наборе параметров особи tr+ действительно практически всегда (10 попыток из 10) вытесняют особей tr-, т.е. «интересы» особи и популяции в данном случае совпали. Однако так бывает не всегда: при некоторых параметрах в модели популяция в целом выигрывает от трансформации, однако аллель tr+ не может в ней закрепиться, поскольку его быстро вытесняет аллель tr-. (...)
Итак, перспективы развития ГГО (трансформации) оказываются менее радужными, если мы посмотрим на проблему с точки зрения индивидуального отбора («интересов особи»), а не группового отбора («интересов популяции»).
Генетический уровень (отбор на уровне генов).
Ситуация усугубляется еще сильнее, если мы вспомним, что до сих пор мы моделировали эволюцию наших микробов не совсем «по-честному». Мы забыли учесть, что сам ген tr тоже может подвергаться трансформации (до сих пор моделировалась только трансформация «генов домашнего хозяйства», а ген трансформации был для трансформации недоступен). Это, конечно, нереалистично.
Что же будет, если мы позволим гену tr+ заменять самого себя на случайно выбранную из генофонда популяции копию гена tr точно так же (с той же частотой), как это проделывают все остальные гены?
Легко увидеть, что трансформация оказывается для аллеля tr+ самоубийственным занятием. Носители этого аллеля будут систематически менять его на tr-, а вот в обратную сторону ГГО работать не будет, потому что носители аллеля tr- не осуществляют трансформацию.
Это резко снижает вероятность фиксации аллеля tr+, вопреки «интересам популяции» и даже вопреки «интересам особи»!
ДЕМОНСТРАЦИЯ: (PS=100, RR=3, NG=20, MR=0.05, BMR=0.1, SD=0.7, TR=0.5)
И интересы популяции, и интересы особи перевешиваются интересами «эгоистичного гена» tr-. Этого оказывается достаточно, чтобы аллель tr+ был вытеснен из генофонда и замещен аллелем tr-.
При этих параметрах аллель tr+ всегда вытесняется аллелем tr-, если ген tr подвержен трансформации, хотя, как мы видели, аллель tr+ при этих параметрах всегда «побеждает» и фиксируется, если он ей не подвержен.
Между прочим, допущение о неподверженности гена tr трансформации является крайне нереалистичным. Бактерии едва ли могут целенаправленно исключить какой-то ген из процесса ГГО, а если и исключат, всегда может появиться в любом другом месте генома новая мутация, препятствующая трансформации. Естественно, вытеснение tr+ и фиксация tr- ведет к снижению Favg, т.е. оказывается вредным для популяции.
ВЫВОДЫ:

ЕСТЬ ЛИ ВЫХОД?
-----------------------------------------

ДЗ: Прочесть 3 раздела, отмеченные галочками. Обратить внимание на примеры (ссылки).

Кто знает, какое отношение имеет эта фотография к теме прошлой лекции?
Почему соотношение полов почти всегда 1:1? (расск.)

Чтобы не «повисла в воздухе» идея эгоистичного гена, приведу один реальный пример (ген tr – это был пример вымышленный, теоретический).
Как правило, распространение аллеля в генофонде популяции определяется той пользой, которую этот аллель приносит организму. Если аллель делает организм более жизнеспособным или плодовитым по сравнению с другими аллелями того же гена, то он будет поддержан отбором и распространится. В этом случае «интересы» гена и организма совпадают, а эгоизм гена никак не проявляется.
Но интересы гена не обязаны всегда совпадать с интересами организма. Если в результате мутации аллель приобретет способность размножаться быстрее других аллелей, принося при этом организму вред, то он всё равно будет поддержан отбором. Другое дело, что это не так-то просто для аллеля – приобрести такую способность (способность размножаться быстрее других аллелей НЕ путем помощи другим генам генома и организму).
Самозарождающиеся в геноме МГЭ; вирусы, происходящие от МГЭ («одичавшие» генов) – это самые простые и очевидные примеры. В этом случае ген приобретает способность размножаться (отчасти) независимо от размножения хозяйского организма. И сразу получает свободу действий (может проявить свой эгоизм).
Но бывает и так, что «эгоистичный ген» не может сам размножаться, но все равно ведет себя эгоистично по отношению к другим генам генома.
Пример. У многих видов в генофонде присутствуют гены (иногда — целые хромосомы), которые при помощи разных замысловатых приемов ухитряются попасть в большее число потомков, чем это предусмотрено законами генетики. Это явление называют «смещением передаваемого соотношения» (TRD), потому что соотношение аллелей (генетических вариантов) у потомства оказывается смещенным относительно ожидаемого, то есть менделевского.
Один из хорошо изученных примеров «эгоистичного гена», вызывающего TRD, — так называемый гаплотип t, встречающийся у домовых мышей (Mus musculus, это еще один классический объект). Это, собственно, не один ген, а комплекс генов, расположенный на 17-й мышиной хромосоме. Из-за нескольких инверсий гаплотип t не рекомбинирует (не обменивается участками во время мейоза) с соответствующим фрагментом «здоровой» 17-й хромосомы. Поэтому он наследуется как единое целое, что дает возможность генам, входящим в его состав, хорошо приспособиться друг к другу и «согласовать» свою коварную деятельность по осуществлению мейотического драйва.
Самцы с генотипом +/t, имеющие одну 17-ю хромосому «дикого типа», а другую — с гаплотипом t (то есть гетерозиготные по гаплотипу t), должны были бы, согласно законам классической генетики, передавать гаплотип t только половине своих потомков. В действительности гаплотип t наследуется большинством (до 99%) потомков такого самца. Это и есть TRD. У самок гаплотип t не вызывает TRD, то есть самки +/t передают гаплотип t ровно половине своих детей.
Как удается гаплотипу t обмануть механизм «честного» (равновероятного) распределения хромосом по гаметам, на котором основаны законы Менделя? Оказалось, что самцы +/t, как и положено, производят сперматозоиды + и t в равных количествах. Однако большинство сперматозоидов с нормальной 17-й хромосомой (+) не могут нормально двигаться: у них плохо работает жгутик.
Почему? Оказалось, что гены, входящие в состав гаплотипа t, обездвиживают сперматозоиды + при помощи системы «яд — противоядие». На ранних этапах формирования сперматозоидов в семенниках самца эти гены обеспечивают синтез «яда», то есть белков, блокирующих работу жгутика. Эти белки попадают во все или в большинство будущих сперматозоидов. На более поздних этапах в сперматозоидах t, и только в них, синтезируется «противоядие». В результате сперматозоиды t сохраняют подвижность (и, следовательно, шанс оплодотворить яйцеклетку), а сперматозоиды + оказываются обездвиженными.
Если бы при этом гаплотип t не снижал приспособленность организма, он бы, скорее всего, быстро зафиксировался, то есть полностью вытеснил аллель + и достиг стопроцентной частоты. Тогда он уже не вызывал бы TRD (некого стало бы обездвиживать), и генетики вряд ли обратили бы на него внимание. В конце концов, мало ли в мышином геноме генов с неясной функцией. Не исключено, что судьба многих «эгоистичных генов» именно так и складывается.
Но гаплотип t не может зафиксироваться, потому что он резко снижает приспособленность мышей. В нём содержатся рецессивные летальные мутации. Особи t/t либо погибают на ранних стадиях эмбрионального развития, либо оказываются бесплодными.
Гибель (или бесплодие) гомозигот t/t означает, что естественный отбор на уровне организмов активно противодействует распространению гаплотипа t. Отбор на уровне сперматозоидов действует в обратном направлении. В результате частота t в генофонде стабилизируется на некотором постоянном уровне.
Но все остальные гены – «против», и они «борются». Полиандрия как способ борьбы с распространением гаплотипа t. Поэтому не 30%, а 5-15%.
предыдущий раздел следующий раздел оглавление