
Проблемы Эволюции |
1968
Теория стабилизирующего отбора
II. ДИНАМИКА ИСТОРИЧЕСКОЙ ИЗМЕНЯЕМОСТИ ПОПУЛЯЦИЙ
1. БИОГЕОЦЕНОЗ КАК АРЕНА ПЕРВИЧНЫХ ЭВОЛЮЦИОННЫХ ПРЕОБРАЗОВАНИЙ
Организмы живут и размножаются всегда в совершенно определенной среде, к которой они в известной мере приспособлены. Для каждого вида организмов внешняя среда характеризуется составом воды или почвы, их химическими и физическими свойствами, почвенной флорой и фауной, растительным покровом, животным населением, а также целым комплексом факторов, которые можно назвать климатом и микроклиматом в широком смысле (влажность почвы и воздуха, температура, освещенность, течение воды и движение воздуха, а также периодические и случайные изменения этих факторов).
Приспособленность означает нормальную жизнедеятельность данного организма в данных условиях существования и характеризуется сложным взаимодействием со всеми указанными факторами, которые таким образом входят в нормальную его жизненную обстановку. Эта приспособленность выражается именно не только по отношению к факторам неорганической среды, но и по отношению к совместно с ним в этой среде обитающим другим организмам. С последними он может конкурировать или сожительствовать, может их использовать в качестве пищевого материала, а может наоборот, служить пищей для других организмов. Таким образом устанавливаются очень сложные связи, которые все вместе образуют нормальные условия существования данного вида организмов. Эти условия различны в разной среде (водной, воздушной, подземной), на разных почвах, в различных местах (биотопах), в различных климатах и микроклиматах, при различном составе растительного и животного населения. Однако всегда ясно выражена взаимная приспособленность организмов в типичных местообитаниях. Организмы образуют взаимосвязанные комплексы, характерные для определенных местообитаний. Такие типичные сообщества организмов называются биоценозами. В состав последних входят, следовательно, как характерный растительный покров, так и животное население. Необходимость этой взаимосвязи понятна уже из того, что все животные существуют в конце концов за счет растений. Однако растения черпают необходимые им жизненные средства из окружающей неорганической среды (из углекислоты воздуха и из воды с растворенными в ней солями почвы). Поэтому, рассматривая жизнь данного комплекса организмов (биоценоза), мы не должны упускать связи с данными почвенными и климатическими факторами, которые являются такой же основой существования растительного покрова, как последний является основой для жизни животного населения. Поэтому мы в дальнейшем будем пользоваться термином В. Сукачева — биогеоценоз — для обозначения взаимосвязанных комплексов организмов, характерных для известных геофизических условий.
В понятие биогеоценоза входят все связи между организмами и средой, а следовательно, как взаимозависимости между разными видами растений и животных, так и взаимозависимости между особями одного и того же вида. Сказанное об определенности взаимоотношений между элементами биогеоценоза вовсе не означает, однако, неизменности его состава. Здесь возможны как случайные, так и периодические колебания, а также исторические изменения, которые тогда неизменно сопровождаются и изменениями в самих организмах. Все эти изменения несомненно регулируются в известной степени действием внутренних сил самого биогеоценоза. Вхождение известных организмов в состав определенных биогеоценозов не означает также их строгой связанности. Некоторые виды организмов могут в разных возрастах (стадиях развития) или в разные сезоны входить в разные биогеоценозы. Некоторые подвижные или вообще эврибионтные организмы могут вообще входить в состав разных биогеоценозов (очень часто у птиц, например,— гнездование в одном биоценозе, а кормление — в другом).
Среди взаимозависимостей между организмами, входящими в состав определенных биогеоценозов, основное значение имеют пищевые связи как прямые (типа взаимоотношений между хищником и жертвой), так и косвенные (пищевая конкуренция).
2. ПОПУЛЯЦИЯ КАК ЭЛЕМЕНТАРНАЯ ЭВОЛЮИРУЮЩАЯ ЕДИНИЦА
Биоценозы, как правило, не имеют сплошного распространения. Они распределяются по биотопам с соответствующими почвенно-климатическими условиями. Соответственно этому и особи определенного вида организмов не имеют сплошного распространения. Входя в состав определенных биогеоценозов, они распределяются группами по подходящим биотопам, образуя в каждом из них более или менее обособленное население — популяцию особей данного вида организмов. Величина и обособленность популяций могут быть весьма различными и это зависит как от пространственных соотношений (большой и однородный лес или маленькая роща, река, озеро или пруд, безграничная степь, луг в пойме реки или лужайка в лесу, большое заболоченное пространство или отдельное болотце и т. п.), так и подвижности самого организма (способ рассеивания семян у растений, активное перемещение у животных). Во всяком случае жизнь и размножение особей данного вида организмов протекает в популяциях разной величины и разной обособленности, входящих в состав определенных биогеоценозов.
Эволюция определяется борьбой за существование в условиях сложных взаимоотношений между элементами биогеоценоза. Эти соотношения не являются постоянными. Кроме случайных и периодических колебаний, можно установить также наличие исторических изменений состава целых биогеоценозов. Наиболее существенным, основным выражением исторического преобразования живых систем (включая биоценозы) является эволюция морфофизиологической организации особей как представителей определенного вида организмов. Эта эволюция протекает только в ряду поколений, т. е. в целых линиях особей, совместно составляющих данную популяцию во временной преемственности ее структуры. Наиболее типично протекает эволюция в популяциях, состоящих из свободно меж собой скрещивающихся особей. Только при скрещивании происходит перекомбинирование наследственных свойств как важнейший процесс перестройки наследственной основы организации.
Возможность свободного скрещивания фактически ограничивается главным образом лишь особями одной популяции, хотя обычно возможен и более или менее свободный обмен мигрантами между разными популяциями одного и того же вида. Как уже сказано, степень обособленности между популяциями может быть весьма различной и это зависит не только от территориальной их близости, но и от степени подвижности и средств расселения данного вида организмов. Соревнование особей и естественный отбор протекают также в основном внутри популяции и если биогеоценоз является ареной первичных эволюционных преобразований, то входящая в его состав популяция данного вида организмов является наименьшей эволюирующей единицей.
До тех пор, пока вид еще не распался на вполне изолированные новые единицы, пока не прекратились обмен мигрантами и внутривидовые скрещивания, он изменяется в процессе эволюции как более или менее сложная, но все же целостная система. При этом руководящее значение имеют сложные взаимоотношения организмов с внешней средой, названные Дарвином борьбой за существование. Эта борьба связана с гибелью (элиминацией) одних, как правило, менее приспособленных, особей и естественным отбором других, имеющих в среднем известные преимущества в данных условиях внешней среды. В результате этого одни формы исчезают из популяции, а другие получают в ней все большее распространение. Состав популяции при этом меняется. В редких случаях удается непосредственно установить изменение численных соотношений различных типов особей, входящих в состав популяции. Так, например, у божьих коровок (Coccinellidae) популяции одного и того же вида состоят из особей различной окраски. Географически различные популяции содержат те же типы, но в различном численном соотношении (Harmonia axyridis, рис. 10, Dobzhansky Т., 1933, 1937). Различное направление естественного отбора ведет к изменению этого соотношения даже в одной и той же изолированной популяции по сезонам. Весной у Adalia bipunctata (рис. 18, табл. 2) преобладает «красная форма», осенью— «черная», которая летом, очевидно, размножается с большей интенсивностью, а зимой гибнет в большем числе (Timofeeff-Rossovsky, 1940).
Географическое изменение численного соотношения между альтернативными типами в популяциях наземных моллюсков рода Partula, населяющих различные долины островов Таити и Муреа, установлено Крэмптоном (Crampton, 1932). Популяции даже соседних долин различаются по комбинациям признаков, охватывающих окраску, величину и форму раковины, правое и левое вращение раковины моллюска.
Гораздо чаще популяции имеют фенотипически однородный характер. Тогда географическая изменяемость представляется первоначально непрерывной по всем признакам. Дж. Гексли (J. Huxley, 1940) говорит в этом случае о «градиенте» признаков (Charakter-gradients, clines). Несмотря на непрерывность связи, географически наиболее удаленные представители такого вида могут быть настолько различны, что не только по морфологическим признакам, но и физиологически (по нескрещиваемости) заслуживают выделения в самостоятельные виды. Много фактов такого рода собрано у Ренша (Rensch, 1929, 1939). Нас, однако, занимает здесь не самая проблема видообразования, а лишь процесс возникновения стойких различий.
Хорошим примером непрерывной географической изменяемости может служить из растений обыкновенный сон-прострел (Anemone Pulsatilla, Zimmermann, 1938). Он дает непрерывный ряд форм, закономерно изменяющихся в направлении с запада на восток Европы. На западе он обладает тонко рассеченными и разбросанными листьями и поникшими цветами. На востоке у него более грубо рассеченные, стоячие листья и стоячие цветы. Эти различия имеют наследственный и вместе с тем приспособительный характер — они связаны с обилием дождей на западе и засушливостью в восточной Европе. Положение листьев способствует стеканию дождевой воды к корневой системе у восточных форм и, наоборот, разбрызгиванию воды у западных. Поникшее положение цветов предохраняет от смывания пыльцы дождевой водой. Степень рассеченности листьев связана с различной интенсивностью транспирации. Мало рассеченные листья восточной формы испаряют значительно меньше воды, чем богато расчлененные листья западной формы (что доказано прямым измерением транспирации в одинаковых условиях опыта). Из растений можно еще привести подобный пример переходов между горной формой люцерны Medicago glutinosa и степной формой Medicago jalcata. На горных лугах Балкарии и Карачая встречается типичная М. glutinosa с опушенными, спирально закрученными плодами (бобами). При переходе на нижележащие луга опушение теряется и плод раскручивается. Medicago glutinosa постепенно переходит в типичные формы М. jalcata, которые в степи, на границе гор, обладают прямыми и голыми плодами (Синская, 1938).
ТАБЛИЦА 2. Число переживающих зиму красных и черных божьих коровок Adalia Bipunctata по результатам подсчета на местах зимовки в предместье Берлина

Примером непрерывной географической изменяемости могут служить улитки Cerion на Багамских островах. На острове Нью-Провидаис они постепенно и закономерно изменяются при переходе с запада на восток. Западные формы — белые и ребристые; восточные — слабо ребристые или почти гладкие и пигментированные. Все они связаны совершенно непрерывными переходами, дающими полный ряд промежуточных форм между обоими крайними типами. Изменения, по-видимому, связаны с различиями в климате — на востоке более дождливом, на западе — засушливом. Более толстая, белая раковина западных форм, очевидно, лучше защищает тело от чрезмерного нагревания на солнце (L. Plate, 1907).
Постепенный ход изменений установлен В. Алпатовым (1927) для домашней пчелы. При переходе с севера на юг можно отметить уменьшение общих размеров, удлинение конечностей и удлинение хоботка. Последнее связано, очевидно, с использованием других цветов, обладающих более глубокими нектарниками. Жук-жужелица Discoptera komarovi дает при биометрической обработке картину непрерывного ряда изменений в направлении с востока Туркестана на запад до Каспия. Закономерно меняется целый ряд признаков. Приспособление к жизни в барханных песках ведет ко все большей специализации западных популяций. Очевидно, эволюция этого жука сопровождалась его постепенным распространением. Восточные популяции D. komarovi филогенетически старше и стоят по своей организации ближе к исходным туркестанским формам. Прикаспийские популяции наиболее молоды, но и наиболее изменены (К. Арнольди, 1939).
Аналогичные исследования, сопровождавшиеся генетическим анализом, были проведены в большом масштабе Р. Гольдшмидтом (R. Goldschmidt, 1927, 1935) над непарным шелкопрядом (Porthetria dispar). Этот вид обладает широким распространением в Европе и Азии и распадается на много географических рас, связанных между собой всеми переходами. Они различаются по общей величине, по пигментации и другим признакам. Они различаются и по своему жизненному циклу, и здесь ясно видна приспособительность расовых различий по отношению к местным условиям климата. Северные и горные популяции, соответственно более краткому периоду вегетации, обладают более быстрым эмбриональным развитием и более коротким периодом личиночного развития, чем южные расы. Здесь, как видно, преобладают физиологические различия.
В других случаях именно физиологические различия выступают на первый план, и географические расы морфологически еще неразличимы. Так, у Drosophila funebris установлено существование географических рас, обнаруживающих приспособительные различия в стойкости по отношению к крайним температурам и в температурном оптимуме. Восточная раса, населяющая СССР, весьма устойчива как к высоким, так и к низким температурам. Северо-западная раса довольно устойчива к низким температурам, в то время как юго-западная раса, распространенная по средиземноморским странам, весьма устойчива по отношению к высоким температурам (Timofeeff-Ressovsky, 1935a). Сходные физиологические различия (по плодовитости в различных условиях температуры) были установлены для различных популяций Drosophila pseudoobscura (Dobzhansky, 1937). Здесь мы имеем, несомненно, лишь самые начальные этапы дифференцировки единого вида на местные географические или экологические формы.
Мы не ставим себе задачей последовательный анализ процессов видообразования. Это делалось многими современными авторами (В. Л. Комаров, 1940; Rensch, 1939; Dobzhansky, 1937; Сборник под ред. J. Huxley — The new systematica, 1940, и др.). Обратим, однако, внимание на следующие факты. Генетический анализ указанных межрасовых различий показал,— во всех случаях, когда такой анализ был возможен, — наличие наследственных изменений (хотя наряду с этим могли быть и модификационные изменения, например, у той же Anemone Pulsatilla). Эти различия имеют дискретный характер мутационных изменений. При скрещивании различных рас они показывают картины менделевского расщепления.
Однако в подавляющем ряде случаев различия касаются огромного числа «генов», т. е. представляют результат комбинирования весьма многочисленых мутаций. Гибриды первого поколения обычно довольно однородны, а во втором поколении обнаруживается значительное увеличение изменчивости по всем признакам. Это указывает на подавляющее значение многочисленных малых мутаций при формировании географических (и экологических) рас.
Уже простые различия в окраске божьих коровок Harmonia ахуridis (от желтых с черными пятнами до черных с желтыми пятнами) определяются изменениями в трех генах (Tan and Li, 1934). По Семнеру (Sumner, 1920, 1932), географические изменения грызунов Peromyscus определяются изменениями очень многих генов, что доказывается сильным возрастанием изменчивости во втором поколении гибридов. Гольдшмидт (R. Goldschmidt, 1927–1935) находит, что различия географических рас непарного шелкопряда Porthetria dispar простираются на «силу» половых факторов, на окраску гусениц, на величину и окраску взрослых бабочек, на длительность зимнего покоя яиц, на скорость развития и другие признаки. Для каждого признака можно установить влияние целого ряда «генов», одни из которых вызывают значительный эффект, а другие модифицируют его в сторону усиления или ослабления.
Многочисленные мелкие наследственные различия установлены и Бауром (Е. Ваш, 1924) при анализе различных природных популяций львиного зева (Antirrhinum). Наконец, и сравнение местных популяций и рас различных видов дрозофилы показало существование многочисленных различий, основанных, главным образом, на различной концентрации одних и тех же генов. Встречаются и более крупные различия, связанные со стерильностью гибридов между такими популяциями (расы А и В у Drosophila pseudoobscura).
В некоторых случаях различия популяций и рас имеют как будто иной характер и определяются изменением одного только гена. В таких случаях говорят о распространении одной простой мутации. Известным примером такого распространения являются мела-нистические хомяки (Cricetus cricetus), изученные сначала зоогеографом Лепехиным, а затем рядом других исследователей; в новейшее время их распространение изучали С. В. Кириков (1934) и С. М. Гершензон (1946). Они концентрируются у нас главным образом в двух очагах — в Башкирии и на Украине. В обоих случаях их ареал ограничивается более влажными зонами лесостепи.
В Западной Германии описана также простая рецессивная мутация полевки (Microtus arvalis), отличающаяся измененным строением коренных зубов. Ее концентрация достигает в Шлезвиг-Гольштейн, очевидном очаге ее распространения, 85% всех особей. По направлению на юг относительная численность мутантов быстро падает (Zimmermann, 1935).
У божьей коровки Epilachna chrysomelina (Coccinellidae) известна полудоминантная мутация Elaterii, которая характеризуется определенным изменением рисунка. Она образовала в части средиземноморской области (Сев. Италия, Южн. Франция, Корсика, Сардиния) хорошо выраженную географическую расу. Генетически она составляется из одной большой мутации и ряда малых.
Это последнее представляет, очевидно, почти общее правило, относящееся ко всем случаям распространения «отдельных», более значительных мутаций.
Исследования межвидовых гибридов хлопчатника (Харланд, 1937) показали, что этим же путем могут устанавливаться и видовые различия, в которых всегда участвуют многочисленные малые наследственные изменения, хотя ведущее значение и могла иметь в свое время единичная мутация. В огромном большинстве случаев ясно выраженные географические расы отличаются весьма значительным числом генных мутаций, большинство которых связано с незначительными количественными изменениями во многих морфологических и физиологических признаках. Нередко можно установить и приспособительный характер различий, п это указывает тогда на ведущую роль естественного отбора в их накоплении. Вместе с тем ясно, что естественный отбор действует всегда на фоне множества мелких различий, имеющихся между особями любой популяции. Концентрация определенных изменений (мутаций) в определенных популяциях и притом не всегда явно приспособительных; изменения этой концентрации по сезонам, по периодам колебания общей численности; существование известных различий в накоплении мутаций в больших и малых популяциях,—все это указывает на существование довольно сложного комплекса факторов, определяющих динамику исторического преобразования популяции. В этом отношении имеют значение прежде всего, конечно, борьба за существование и естественный отбор, но кроме того — также комбинирование в процессах скрещивания и изоляция, т. е. ограничение этого скрещивания. Сверх того мы должны отметить значение и некоторых менее явных процессов развития системы регуляторных механизмов формообразования и накопления мутаций в скрытом виде, что приводит к внешне незаметной перестройке генотипа. Наконец, мы подчеркнем и значение индивидуальной приспособляемости, основанной на широком развитии системы реакционных механизмов.
3. БОРЬБА ЗА СУЩЕСТВОВАНИЕ И ИНТЕНСИВНОСТЬ ЭЛИМИНАЦИИ
В основе дарвиновской теории естественного отбора лежит борьба за существование, с необходимостью вытекающая из безграничного стремления организмов к размножению. Это стремление выражается всегда в геометрических прогрессиях. Дарвин ссылается при этом на Мальтуса. Однако еще задолго до Мальтуса биологи учитывали потенциальную интенсивность размножения животных и растений. Еще К. Линней указывал, что одна мясная муха при посредстве своего потомства могла бы в немного дней уничтожить до костей труп лошади или льва. Еще в начале прошлого века К. Эренберг подсчитал, что одна диатомея могла бы в восемь дней дать массу материи, равную объему Земли, а в течение следующего часа могла бы удвоить эту массу. Еще быстрее идет в благоприятных условиях размножение бактерий. Однако даже наиболее медленно размножающиеся животные — современные слоны — могли бы, по подсчету Дарвина, очень быстро овладеть всей поверхностью суши, если бы для этого оказались подходящие условия. По Дарвину, от одной пары слонов за 740–750 лет получилось бы около 19 миллионов особей. За 3000 лет слоны покрыли бы сплошным покровом всю поверхность земного шара.
Здесь следует учитывать, что проявления жизненной энергии организмов, как деятелей природы, далеко не исчерпываются одной лишь способностью их к размножению. Плодовитость организмов вовсе не является верным мерилом их реальной активности. Скорее наоборот — плодовитость организмов прямо определяется их потребляемостью, т. е. указывает на малую их приспособленность и на большую активность их врагов. Наконец, мы здесь особо отмечаем, что хотя стремление к беспредельному размножению и лежит в основе эволюции органических форм, однако и темп эволюции не определяется плодовитостью, т. е. показателем геометрической прогрессии размножения. И в данном случае скорее наоборот — решающее значение имеют иные формы активности организма.
Размножаемость потенциальная и реальная. Расхождение между потенциальной и реальной размножаемостью может иногда быть весьма значительным, в особенности при быстрой смене поколений. Как раз в этом случае иногда возможна и реализация геометрической прогрессии размножения. Однако это происходит всегда лишь как местное и притом временное явление. Как правильно отметил Дарвин, «многочисленность яиц представляет значение для тех видов, которые зависят от колеблющихся количеств пищи, так как позволяет им быстро возрастать в числе. Но настоящее значение многочисленности яиц или семян заключается в том, чтобы покрывать значительную их убыль, вызываемую истреблением в каком-нибудь периоде их жизни, а этот период, в большей части случаев, бывает очень ранний» ... «во всяком случае, среднее число животных или растений зависит только косвенно от числа яиц или семян».
В исторической перспективе эволюционного процесса для нас имеют наибольшее значение именно эти средние числа и их стойкие изменения в процессе эволюции. У медленно размножающихся животных и растений (а сюда относятся как раз высшие, прогрессивные их представители) мы имеем дело практически всегда с этими лишь немного колеблющимися средними численностями. Эти средние численности конкретных популяций устанавливаются в результате реализации некоторых средних показателей размножения и некоторого относительно постоянного уровня смертности. Ясно, что такие формы со сравнительно стабильной численностью оказывают и некоторое довольно постоянное давление на окружающую среду.
Реальная размножаемость организма определяется его плодовитостью, т. е. числом зачатков, воспроизводимых в течение всей его жизни. При половом размножении животных это будет число откладываемых яиц или число рождаемых детенышей. У высших растений — число созревающих семян. Это число, конечно, не константно, оно меняется в зависимости от условий данного года или сезона, а также местности с ее жизненными ресурсами. Однако в общем оно колеблется около некоторого среднего уровня. Случайно благоприятные условия, необычные условия известного сезона или периодические изменения в количестве кормового материала или в числе врагов и паразитов приводят к изменениям плодовитости и в особенности к изменению истребляемости яиц, зародышей (семян), молоди и зрелых особей. Это, в свою очередь, ведет к более или менее резким временным изменениям численности, которые соответственно могут иметь характер случайных вспышек, вполне закономерных сезонных колебаний или более или менее правильных периодических явлений усиленного размножения. В этих случаях проявление жизненной энергии организма, как представителя известного вида, его давление на окружающую среду возрастает до известного максимума, после чего оно вновь падает до некоторого минимума проявления жизни вплоть до нового подъема воспроизводительной деятельности. Существование таких колебаний нисколько не меняет наших рассуждений, так как в процессе эволюции имеет значение не эта периодика жизненных проявлений, а более постоянная энергетическая характеристика организма, выражающаяся в средних числах. Эти же средние числа имеют гораздо более стабильный характер и неплохо характеризуют современное положение организма в его нормальной внешней среде.
Интенсивность реального размножения организма определяется его плодовитостью, т. е. числом откладываемых яиц (при половом размножении) и временем развития организма от яйца до наступления половой зрелости. Последнее определяет скорость смены поколений и играет огромную роль в геометрической прогрессии размножения малых организмов (при благоприятных условиях). Именно быстрая смена поколений позволяет организму максимально использовать создающиеся на время благоприятные местные условия существования (использование трупа бактериями или личинками мухи, использование живого организма размножающимися в нем паразитами, использование благоприятных условий данного сезона и т. п.). Более устойчивые формы размножения, интересующие нас в связи с вопросом о борьбе за существование как базе эволюции, определяются в основном плодовитостью организмов.
Именно эта плодовитость организма и характеризует реальную (а не потенциальную) его размножаемость как одно из проявлений «давления жизни». В самом деле, если средняя численность организма в данное время остается вообще довольно постоянной, то это означает, что им ежегодно воспроизводится именно столько новых жизней, сколько ежегодно гибнет от различных причин. Число размножающихся особей не увеличивается, и поэтому «давление жизни», реально проявляемое определенным организмом, остается в общем постоянным. Если N особей производит ежегодно п яиц или молоди, то такое же число п особей различного возраста (главным образом молоди) ежегодно гибнет. Число особей (данной популяции) (N+n—n=N) остается постоянным. Геометрическая прогрессия не определяет реального «давления жизни», так как неродившиеся и погибшие особи фактически не оставляют никакого потомства. Реальное «давление жизни» определяется только фактически существующими особями любого возраста, а не теми, которые могли бы родиться, если бы были подходящие условия.
Размножаемость как мера потребляемости. Если число нарождающихся жизней в среднем равно числу гибнущих, то мы получаем вполне объективный показатель интенсивности борьбы за существование. Плодовитость организма (число яиц, откладываемых животным за всю жизнь, или число семян, производимых растением), отнесенная к единице времени (году), может служить таким мерилом общей интенсивности борьбы за существование, т. е. суммарной элиминации организма во всех его возрастах (начиная со стадии яйца и до зрелого состояния).
Организм в своем стремлении к безграничному размножению встречает известное противодействие среды. Чем выше фактическая размножаемость, тем сильнее это противодействие, выражающееся в истребляемости (элиминации) организма. Это понятно, так как размножаемость организма, при среднем постоянстве его численности, равна его истребляемости. Это условное подвижное равновесие поддерживается на известном уровне благодаря своеобразию взаимоотношений между организмом и средой и в особенности благодаря теснейшей взаимозависимости между различными организмами. Всякое увеличение средней численности какого-либо вида организмов обостряет конкуренцию между особями и при нехватке света (для растений), пищевых ресурсов, воды, кислорода и других жизненных средств ведет к ухудшению жизненных условий, к снижению плодовитости, к повышению смертности и, следовательно, к уменьшению численности особей. Уменьшение численности ниже обычного уровня ведет, наоборот, к смягчению внутригрупповой конкуренции, к улучшению жизненных условий и к повышению плодовитости, а, следовательно, к увеличению числа особей данной популяции. Увеличение числа особей означает также улучшение условий существования для хищников и паразитов данной формы, стимулирует их размножение, что, в свою очередь, ведет к повышенной истребляемости преследуемой формы. К этому прибавляется еще ускоренное распространение инфекционных заболеваний при достижении известной плотности населения. Развивающиеся эпизоотии вызывают массовый падеж, резко снижающий численность чрезмерно размножившейся формы.
Все эти и другие подобные же факторы играют, таким образом, роль регуляторов плотности населения любого вида животных или растений, поддерживающих его численность в среднем на известном нормальном для данной эпохи уровне.
Нас, однако, здесь не интересует биологическое равновесие форм в окружающей нас природе. Мы воспользовались лишь его бесспорным наличием, в масштабе времени наших обычных наблюдений, для установления возможности довольно точного измерения общей интенсивности борьбы за существование любого вида организмов. К сожалению, этот метод — учет реальной размножаемости — не дает возможности дифференцированного подхода. Мы учитываем лишь суммарную истребляемость организма во всех его возрастах и не можем воспользоваться этим косвенным методом для учета интенсивности элиминации на различных стадиях развития. Для этого необходимо непосредственное определение истребляемости данной формы. Однако в некоторых случаях и величины суммарной элиминации могут дать достаточно верное представление не только об интенсивности, но и о характере борьбы за существование, в особенности при возможности сравнительного изучения плодовитости и экологии многих близких форм.
Так, например, из двух видов черноморской улитки Nassa один вид iV. reticulata откладывает в год свыше 22 000 яиц, другой, более специализированный вид N. neritea откладывает, также в год, около 10–15 яиц. Так как это сходные виды, ведущие одинаковый образ жизни, но различающиеся по своему развитию,— N. reticulata проходит стадию пелагической личинки veliger, a N. neritea обладает прямым развитием,— то ясно, что у N. reticulata огромная истребляемость падает на время свободной личиночной жизни в планктоне (М. Бекман, 1941).
Нас интересует здесь, как сказано, не наличие равновесия, а возможность его стойкого смещения в связи с процессом эволюции. Поскольку в основе последней лежит борьба за существование, измерение ее интенсивности приобретает первостепенное значение. Борьба за существование ведет непосредственно к естественному отбору. Однако определяется ли интенсивность отбора интенсивностью борьбы за существование, т. е. элиминации, и не связано ли направление отбора с различными формами этой элиминации?
Размножение организмов и противодействие внешней среды этому размножению. Противодействие среды размножению организма выражается в различных формах, ведущих, однако, к одному и тому же конечному результату — к истребляемости, т. е. прямому уничтожению или устранению от размножения известного числа особей. Роль среды выражается столь различно, что этого нельзя не учитывать. Менее всего она исчерпывается обычным для экологов понятием сопротивления. И среда, включающая всю органическую природу, с которой организм вступает в ту ила иную связь, далеко не только «сопротивляется» размножению, она и сама ведет наступление на организм, независимо от темпов его размножения. Поэтому можно различать, кроме депрессивного, еще и агрессивное противодействие среды. Между этими выражениями деятельности среды имеются коренные различия, которым соответствуют различное положение организма в этой среде и, следовательно, различные формы борьбы за существование.
Депрессивная роль среды. Если предел численности данного вида организмов диктуется главным образом ограниченностью жизненных средств и, в первую очередь, ограниченностью продуктов питания, то размножение организма наталкивается именно на сопротивление среды; оно насильственно подавляется недостатком пищевого материала и наступающей вследствие этого физиологической депрессией. Обострение активной конкуренции, связанной с косвенной элиминацией многих особей данного вида, ведет к уменьшению плодовитости и, через естественный отбор на максимальную экономичность (Шмальгаузен, 1939а),— к прогрессивной специализации.
Так как ограниченность жизненных средств, при отсутствии высших форм заботы о потомстве, отзывается в особенности тяжело на развивающейся и растущей еще молоди, то она нередко ведет к увеличению размеров детенышей, а, следовательно, и к возраста-ниго окончательной величины взрослого животного. В этом увеличении объема яиц, величины детенышей и общих размеров тела мы видим наиболее простую, но далеко еще не совершенную форму максимального обеспечения своего потомства. Депрессивное ограничение жизненной энергии организмов ведет их по пути теломорфоза (И. Шмальгаузен, 1939а), завершающего прогрессивную эволюцию большинства филогенетических ветвей. Такова судьба в особенности наиболее защищенных организмов, занимающих высшие ступени филогенетической лестницы. Именно защищенность от врагов ведет к усиленному размножению и обострению внутри-групповой конкуренции с ее последствиями. В особенности это касается хищников, занимающих места высших звеньев в цепях питания (пищевых рядах). Здесь специализация питания и увеличение общих размеров становятся ведущими процессами эволюции.
Агрессивная роль среды. Если предел численности данного вида организмов диктуется не нехваткой жизненных средств, а его систематическим истреблением другими организмами, для которых он служит пищей, то лишь с большой натяжкой можно говорить о сопротивлении среды. Размножение осуществляется, в сущности, не только совершенно свободно, Но даже поощряется. Все преимущества в борьбе за существование находятся теперь на стороне наиболее плодовитых особей. Среда в лице хищников и паразитов не «давит», а неудержимо поглощает свой пищевой материал, предоставляя для остающихся в я^ивых явный избыток жизненных средств и неограниченные возможности размножения. Преобладание пассивных форм конкуренции и прямой элиминации ведет к повышению плодовитости, к раннему созреванию, к выработке средств защиты и, при очень интенсивной элиминации, к прогрессивному измельчанию, как наиболее простому, весьма несовершенному еще способу защиты своей жизни, а также к известному недоразвитию, к деспециализации. Таким образом, агрессивное ограничение жизненной энергии организмов ведет их нередко по пути катаморфоза (и гипормофоза, И. Шмальгаузен, 1939а). Такова судьба наименее защищенных организмов, занимающих обычно низшие ступени филогенетической лестницы. В экономии природы они располагаются на низших звеньях цепей питания (пищевых рядов). Здесь известная деспециализация, измельчание и высокая плодовитость приобретают ведущее значение в процессе эволюции.
Можно было бы иллюстрировать эти пути эволюции многими конкретными примерами из области экологии или еще более убедительными данными палеонтологии. Увеличение размеров и специализация характерны всегда для наиболее защищенных во взрослом состояние организмов данной эпохи. Это показывает история стегоцефалов в каменном угле и в перми, история рептилий в мезозое и млекопитающих в третичном периоде. После появления новых, более совершенных, более активных форм специализированные формы вымирают, а остающиеся мельчают — такова позднейшая судьба амфибий, начиная с триаса, судьба рептилий в третичный период. Интересно, что даже паукообразные и насекомые сначала, когда они одни еще царили на суше, пошли по пути увеличения размеров тела (каменноугольные Palaeodictyoptera, прямокрылые и стрекозы с размахом крыльев до 75 см). С появлением, однако, подвижных наземных позвоночных (рептилий и затем птиц) они не только перестали увеличиваться в размере, но определенно пошли по пути измельчания. В особенности это касается клещей и малоподвижных насекомых. В мезозое хищные динозавры питались, очевидно, между прочим и млекопитающими, а млекопитающие — яйцами и молодью динозавров. Это привело к дальнейшей специализации динозавров и к утере ими пластичности. Вместе с тем это решило судьбу небольших, но активных и плодовитых млекопитающих.
Формы активности организма в его борьбе за существование. Если мы отмечаем значение различных форм активности среды, противодействующей безграничному стремлению организмов к размножению и расселению, то еще большее значение имеют в процессе эволюции различные формы активности самого организма в его борьбе за существование.
Известная активность является одной из наиболее общих характеристик всего живого, и если мы только что говорили об активности внешней среды, то в ее основе лежит ведь именно активность повсюду населяющих ее живых организмов. Формы и степени этой активности весьма разнообразны. Активность какого-либо лишайника в использовании минеральных веществ, в борьбе за свет и воду, конечно, неизмеримо ниже, чем активность птицы или млекопитающего, поражающих нас своей непрерывной деятельностью. И все же в работе лишайника и в поведении млекопитающего можно найти немало сходства. Вся эта активность так или иначе направлена на сохранение жизни. В первую очередь она выражается в захватывании необходимой материи и энергии, во вторую очередь — в защите от покушений со стороны других организмов, от их попыток воспользоваться готовыми материалами для построения собственного тела. Эти попытки повторялись в истории организмов многократно. Они привели еще на заре органической жизни к глубокому расхождению между миром растений и миром животных. Растения являются трансформаторами и основными аккумуляторами солнечной энергии. Животные используют готовые растительные материалы и проявляют разнообразнейшую деятельность, на основе дальнейшей трансформации той же солнечной энергии. В дальнейшей истории животных постоянно происходила их пищевая дифференциация, связанная с разделением любой группы на растительноядные и плотоядные организмы, на хищников и паразитов различных порядков. С этой дифференциацией связано и развитие различных форм активности организмов. На первый взгляд кажется, что в процессе эволюции происходит непрерывное в общем его течении возрастание значения особи и что вся деятельность организма направлена на сохранение индивидуальной жизни, и только через это — на поддержку существования данного вида. Это — обманчивое впечатление. На самом деле вся активность организмов направлена только на сохранение преемственности жизни, на обеспечение жизни в потомстве, и само образование более или менее стойкой индивидуальности есть-лишь средство для обеспечения этой преемственности. Поэтому в размножении мы имеем основное и наиболее примитивное проявление активности организмов. При возрастающей потребляемости новым ответом организма будет именно повышение его плодовитости. Уже рост, как другое очень примитивное проявление жизнедеятельности, занимает вполне подчиненное положение. Увеличение органической массы есть необходимая предпосылка для возможности ее раздробления в процессе воспроизведения дочернего поколения. Развитие более крупной и более стойкой индивидуальности связано с продукцией более значительного, более обеспеченного предварительными запасами и потому сразу же более стойкого потомства. Увеличение размеров молоди, связанное с уменьшением ее численности, а нередко также с увеличением размеров зрелой особи, представляет наиболее примитивную форму обеспечения жизни потомства. При возрастающей потребляемости молоди организм, не обладающий иными средствами защиты своего потомства, чаще всего реагирует именно увеличением ее размеров. Продление индивидуальной жизни и периода размножения вместе с увеличением размеров тела является вторым, более совершенным (по сравнению с повышением плодовитости) ответом организма на высокую потребляемость его потомства. Этот ответ характерен для более дифференцированных и стойких форм жизни, обладающих более совершенными средствами защиты. Нередкое в филогенетических ветвях увеличение размеров тела, при переходе от более древних представителей группы к более молодым, иллюстрирует этот путь эволюции форм защиты жизни в своем потомстве. Однако в общем процессе эволюции органических форм и этот довольно примитивный путь оказался несовершенным — он связан с утерей пластичности, с невозможностью быстрой перестройки организации в процессе смены условий внешней среды, с опасностью вымирания. Можно, применяя термин В. О. Ковалевского, говорить о возрастании общих размеров тела и, в особенности, о выраженном гигантизме как об «инадаптивном» пути обеспечения жизни потомства, заводящем организм в тупик эволюции.
В процессе эволюции более сложных организмов неизменно выдвигались вперед более активные организмы, обладавшие и более совершенными формами обеспечения жизни своего потомства, формами, которые оказались гораздо более «адаптивными». Выдвигались вперед те растения, активность которых выражалась в более интенсивном росте и в обеспечении семян средствами быстрого развития. Выдвигались в особенности и более активные животные, действующие как гораздо более быстрые и потому более эффективные трансформаторы энергии. Они быстрее развивались и созревали, быстрее накапливали запасы питательных веществ в многочисленных яйцах, и потому не только оказались опасными конкурентами крупным и малоподвижным специализированным формам прошедшей эпохи, но и обладали, при быстрой смене поколений, исключительной пластичностью при любой смене условий существования. Пассивные средства защиты своего потомства в виде различного рода яйцевых оболочек; инстинкты, заставляющие животное откладывать яйца в скрытые места; защитная окраска и, наконец, охрана своей кладки — все это способствовало более эффективной защите жизни потомства. Однако в процессе дальнейшей эволюции вперед выдвинулись гораздо более совершенные формы заботы о потомстве и в этом, наконец, были достигнуты высшие формы активности самого организма.
Активная деятельность организма в добыче пропитания и защите своей жизни направляется теперь не только косвенно, но и все более непосредственно на обеспечение жизни своего потомства. Организм активно защищает свое потомство от врагов, снабжает его питательным материалом и лишь постепенно вводит его в круг навыков самостоятельной жизни. Вся сложная деятельность птиц и млекопитающих подчиняется в конце концов интересам их потомства и выливается, наконец, в форме вполне организованной жизни некоторой, хотя бы и временной, семьи. Именно с этим связана эволюция высших форм жизнедеятельности организмов, как она реализуется лишь у высших животных.
Высокая активность организма означает высокий темп жизни. Именно эти высокие темпы жизни обеспечили победу высших животных в борьбе за существование. Те же высокие темпы жизни, связанные со значительной пластичностью организмов и, в первую очередь, с быстрой сменой поколений, будучи результатом процесса эволюции, обеспечили и в дальнейшем высокие темпы прогрессивной эволюции.
Уже из этого краткого обзора мы видим, сколь разнообразны формы проявления борьбы за существование и сколь различно их значение в процессе эволюции органического мира.
4. ФОРМЫ БОРЬБЫ ЗА СУЩЕСТВОВАНИЕ
Понятие «борьба за существование» может иметь смысл и оправдание только в дарвиновском широком «метафорическом» понимании: «включая сюда зависимость одного существа от другого, а также включая (что еще важнее) не только жизнь одной особи, но и успех ее в оставлении после себя потомства», «Про двух животных из рода Canis, в период голода, можно совершенно верно сказать, что они борются друг с другом за пищу и жизнь. Но и про растение на окраине пустыни также говорят, что оно ведет борьбу за жизнь против засухи, хотя правильнее было бы сказать, что оно зависит от влажности. Про растение, ежегодно производящее тысячу семян, из которых в среднем вызревает лишь одно, еще вернее можно сказать, что оно борется с растениями того же рода и других, уже покрывающими почву. Омела зависит от яблони и еще нескольких деревьев, но было бы натяжкой говорить о ее борьбе с ними только потому, что если слишком много этих паразитов вырастет на одном дереве, оно захиреет и погибнет. Но про несколько сеянок омелы, растущих на одной и той же ветви, можно совершенно верно сказать, что они ведут борьбу друг с другом. Так как омела рассеивается птицами, ее существование зависит от них, и, выражаясь метафорически, можно сказать, что она борется с другими растениями, приносящими плоды, тем, что привлекает птиц пожирать ее плоды и таким образом разносить ее семена. Во всех этих значениях, нечувствительно переходящих одно в другое, я ради удобства прибегаю в общему термину борьба за существование» (Дарвин).
В этой цитате, так же как и в дальнейшем тексте «Происхождения видов», ясно отражается разнообразие форм борьбы в дарвиновском понимании, и вместе с тем совершенно очевидно выступает значение элементов соревнования во всех формах этой борьбы. Трудность расчленения понятия «борьба за существование» не только в том, что различные его формы в конкретных случаях сложнейшим образом переплетаются и связываются, а также незаметно переходят одна в другую и сменяют друг друга во времени (по возрастам, по сезонам и в историческом процессе изменений организма и среды). Трудность состоит еще и в том, что каждая конкретная форма борьбы за существование может быть независимо охарактеризована с двух различных сторон ее проявления и обе эти характеристики оказываются необходимым дополнением друг друга.
Термином «борьба» Дарвин правильно подчеркнул известную активность организма в его соотношениях со средой, из которой он добывает необходимые средства к жизни. В этой борьбе за жизнь и за размножение множество особей гибнет от самых разнообразных причин: от неблагоприятных физических влияний, от других организмов (хищников и паразитов), а также и от голода, в особенности в неблагоприятное для прокормления время года. Так как при прочих равных условиях гибнут или остаются без потомства всегда более слабые и менее приспособленные особи, то борьба за существование ведет, таким образом, к естественному отбору или переживанию наиболее приспособленных. Борьба за существование всегда связана с уничтожением или устранением от размножения. К. А. Тимирязев, подчеркивавший важное значение этой характеристики борьбы, употребляет для этого термин, предложенный еще О. Контом,— «элиминация». Позднее и независимо от них тот же термин употреблял Л. ""»Морган и затем Л. Плате.
Другая, не менее обязательная, характеристика борьбы за существование определяется элементами соревнования. Это обстоятельство подчеркивалось Ч. Дарвином, но совершенно не отражено в предложенном им термине. Между тем борьба отдельной особи не имеет в процессе эволюции никакого значения. Только в том случае, когда известное число особей одного вида подвергается одной и той же опасности (все равно — абиотической или биотической), одним и тем же ограничениям в своем существовании или в своем размножении, возможен естественный отбор некоторых особей, избегнувших этой опасности или преодолевших эти ограничения. Всегда здесь имеются элементы соревнования между особями одного вида за сохранение жизни, за размножение, в борьбе их за пищу, с врагами и паразитами, с жесткими условиями климата, с голодом и болезнями.
Многие авторы, и прежде всего Ф. Энгельс, указывали на необходимость расчленения понятия борьбы за существование. Такая попытка была сделана Ллойд Морганом и продолжена Л. Плате. Следуя этим авторам, обычно различают: 1) конституциональную борьбу с факторами неорганической среды и болезнями, 2) межвидовую борьбу с другими организмами (с хищниками, паразитами и бактериальными болезнями) и 3) внутривидовую борьбу с себе подобными за пищу, пространство и размножение.
Внутри этих групп проводилось и дальнейшее расчленение. Эта система, однако, логически не выдержана, так как в ней переплетаются оба указанных мною выражения борьбы за существование. Поэтому трудно связать с приведенными формами борьбы определенные формы естественного отбора.
Более плодотворной должна оказаться попытка рассмотреть оба выражения борьбы за существование в отдельности: 1) формы соревнования между организмами, борющимися за жизненные средства и условия размножения, и 2) формы элиминации, т. е. истребления организмов или их устранения от размножения.
А. СОРЕВНОВАНИЕ
В различных формах соревнования подчеркивается роль самого орган измай его индивидуальных особенностей в борьбе за существование. Отмечается значение его средств защиты, его активности в добывании необходимых жизненных средств и его стремления к размножению. При этом дается как бы положительная характеристика борьбы за существование. Различные формы соревнования являются, в такой же мере, как и элиминация, обязательной характеристикой дарвиновского понимания борьбы за существование. Организмы соревнуются между собой в борьбе за одни и те же жизненные средства и за возможность размножения. Следовательно, к этой типичной форме конкуренции относится, между прочим, и дарвиновская борьба самцов из-за самок. Соревнование имеет при этом явно активный характер, и его интенсивность увеличивается с увеличением плотности населения.
Организмы соревнуются между собой, однако, также и в средствах защиты от неблагоприятных влияний климата, в средствах защиты от общих врагов — хищников или паразитов — и в охране своего потомства. В этом случае соревнование имеет пассивный характер, и его интенсивность не увеличивается с увеличением численности населения.
Следует различать внутригрупповое (индивидуальное и межсемейное) соревнование и межгрупповое соревнование. Обе формы могут иметь как активный, так и пассивный характер.
а. Внутригрупповое соревнование протекает на основе индивидуальных различий между особями одной популяции в борьбе их за средства к жизни и размножение (активная форма) и за сохранение своей жизни и жизни потомства, т. е. за средства защиты (пассивная форма).
1. Индивидуальное соревнование совпадает и своей активной форме с обычным пониманием конкуренции за средства к жизни и за размножение (отбор идет при этом на базе косвенной элиминации физиологическими факторами). Соревнование в добыче пищевого материала, обостряющееся при увеличении плотности населения или при сезонной нехватке продуктов питания (зимний голод), ведет к недоеданию, а следовательно, к ослаблению сопротивляемости организма вредным влияниям и к снижению его плодовитости.
Естественный отбор идет по линии специализации питания, развития наиболее экономного обмена веществ на основе наиболее гармоничной организации, а также по линии продления жизни (и размножения), сокращения плодовитости (в смысле уменьшения величины кладок или пометов) и обеспечения развития немногочисленного потомства запасами питательного материала. Специализация и сокращение плодовитости будут основными результатами активного соревнования. Активная конкуренция особей может выражаться и в половой конкуренции за спаривание. Она связана тогда с половым отбором и ведет к развитию некоторых вторичных половых признаков.
В своей пассивной форме индивидуальное соревнование охватывает явления борьбы с неблагоприятными физическими (климат) и биологическими (хищники, паразиты) факторами за сохранение своей жизни и жизни потомства. Соревнование в этом случае не задевает интересов других особей той же популяции и не обостряется по мере увеличения их численности.
Естественный отбор идет при этом (на базе прямой элиминации) в направлении выработки наиболее эффективных средств активной и пассивной защиты от врагов, болезней, холода и других вредных влияний. Большое значение имеет, однако, и крепкая конституция, общая активность организма, развитие его органов чувств и инстинктов.
Индивидуальное внутригрупповое соревнование ведет, следовательно, к преобразованию организации и к развитию приспособлений к данным условиям существования. В этой форме соревнования естественный отбор полностью выявляет свое творческое значение. Можно говорить при этом о формообразовательной, т. е. морфогенной, роли1 индивидуального соревнования.
2. Межсемейное соревнование протекает на основе наследственных различий между членами различных семей. Это не предполагает обязательно существования организованной семьи, так как уже факт известного наследственного сходства ставит все потомство одной пары в сходное положение по отношению к факторам внешней среды. Однако наличие инстинктов, связывающих членов одной семьи (колонии муравьев, пчел), выдвигает эти формы борьбы за существование на более видное место.
Межсемейное соревнование ведет к дальнейшему развитию инстинктов и признаков организации, связанных с жизнью семьи как целого. При этом нередко возникают явления полиморфизма и сложные формы заботы о потомстве. При большой интенсивности активного соревнования за жизненные средства естественный отбор ведет к снижению плодовитости, компенсируемому максимальной защитой яиц и развивающейся молоди.
б. Межгрупповое соревнование протекает (на основе групповых различий) между особями как членами различных популяций (связанных общностью происхождения и совместным размножением, т. е. известной наследственной однородностью) в их борьбе за средства к жизни и размножению, за сохранение своей жизни и жизни потомства. Условием для реализации этой формы соревнования внутри известного вида является обязательно разбивка этого вида на отдельные, хотя бы лишь частично изолированные популяции. Изоляция может быть топографической (географической) с разбивкой по стациям, биологической с распределением по биотопам (экологическая изоляция) или с разделением по времени размножения (сезонная изоляция). Кроме того, она может быть основана на начинающемся уже морфо-физиологическом расхождении (в строении полового аппарата, в инстинктах или в структуре половых клеток), связанном с частичным бесплодием (физиологическая изоляция). Последняя знаменует важнейший этап в обособлении новых форм — становление вида. После этого скрещивание между физиологически обособленными видами уже невозможно, и другие формы изоляции теряют свое значение.
В своей активной форме межгрупповое соревнование означает конкуренцию между особями разных популяций за жизненные средства, за размножение и за условия для размножения (нерестилища, места гнездования и т. п.) Такая конкуренция между разными популяциями связана с прямым или косвенным устранением от размножения и приводит всегда к вытеснению одних популяций другими, более приспособленными к данным локальным условиям. Это не означает обязательно уничтожения целых экотипов, подвидов или видов, так как популяция, вытесняемая в одной местности, либо в одних экологических условиях, может оказаться победительницей в другой местности или в иной обстановке.
В своей пассивной форме межгрупповое соревнование охватывает явления прямой борьбы особей различных популяций за сохранение своей жизни и жизни своего потомства.
Естественный отбор особей, более защищенных от вредных влияний данного климата или от истребления врагами (хищниками и паразитами), ведет к постепенному вытеснению одной популяции другой (в среднем, более защищенной). Поскольку элиминирующие факторы имеют в различных местностях или в различных биотопах несколько иной характер, популяция, вытесняемая в одной местности или в одном биотопе, может получить распространение в другой местности или в других экологических условиях. Истребление целых популяций ведет к углублению разрывов между таксономическими единицами.
Межгрупповое соревнование связано, таким образом с вытеснением одних форм другими на известных территориях и ведет, следовательно, к распределению конкретных популяций по различным стациям и биотопам. Борьба за существование и естественный отбор выступают здесь в роли распределяющего фактора. В этом случае можно говорить о хорогенном значении межгруппового соревнования.
Б. ЭЛИМИНАЦИЯ
В прямом или косвенном истреблении особей и в их устранении от размножения выражается как бы отрицательная сторона борьбы за существование. В формах элиминации отмечается значение как физических, так и биотических факторов среды, прямо уничтожающих множество особей или лишающих их необходимых средств к существованию.
Борьба за существование всегда связана с элиминацией менее приспособленных особей и только через эту элиминацию ведет к естественному отбору, т. е. переживанию и размножению особей, наиболее соответствующих данным условиям существования. Элиминация является обязательным выражением всех форм борьбы за существование. Поэтому различные формы последней связаны и с различными формами естественного отбора как обратной стороны элиминации.
Формы элиминации определяются как элиминирующими факторами (физическими, физиологическими и биологическими), так и объектами элиминации (отдельные особи, целые семьи или целые популяции), их возрастом (возрастная элиминация) и, наконец, характером самой элиминации (стихийная элиминация, постоянная, периодическая, сезонная). Кроме того, особняком стоит элиминация от различных случайных причин. Такая элиминация может иметь массовый характер и наблюдается вообще нередко в очень большом масштабе. Я обозначаю ее термином «общая элиминация». В остальном наибольшее значение имеют индивидуальная элиминация и затем семейная и групповая.
а. При общей элиминации (постоянной или периодической) само переживание отдельных особей не определяется какими-либо их преимуществами перед особями уничтожаемыми. Гибель последних есть дело случая и может зависеть от самых различных причин как физических, так и биологических. Хотя эта форма элиминации как будто не имеет избирательного значения, она все же приводит к отбору на большую плодовитость в силу чисто статистической закономерности. При всяком случайном истреблении всегда имеют больше шансов оставить какое-либо потомство те особи, которые размножаются более энергично, т. е. более плодовитые и раньше созревающие. При случайной истребляемости всегда оказывается более вероятным, что уцелеют хотя бы единичные экземпляры из тысяч потомков одной особи, чем из какого-либо десятка или сотни потомков другой особи, которые могут быть полностью уничтожены. В таком случае элиминация получает до известной степени характер семейной элиминации.
Таким образом, общая элиминация означает естественный отбор на максимальную плодовитость. Высокая плодовитость многих мелких рачков и других массовых форм планктона, а также многих насекомых есть результат, а вместе с тем и показатель их высокой общей элиминации (истребление рыбами, птицами и млекопитающими). Особенно интенсивна, однако, массовая гибель яиц и личинок, а вместе с тем и плодовитость многих паразитов. Из растений велика гибель спор и мелких семян у форм, распространяемых ветром и не обладающих значительным запасом питательных веществ в семенах.
б. При индивидуальной элиминации (физическими, физиологическими или биологическими факторами) особи, выживающие и оставляющие потомство, в среднем отличаются по морфологическим и физиологическим признакам от особей истребляемых. Элиминирующие факторы могут действовать с разной интенсивностью на различные возрасты (возрастная индивидуальная элиминация), могут изменяться по сезонам, а также в разное время дня (периодическая элиминация) и в разные фазы жизнедеятельности животного. Все это накладывает свой особый отпечаток на борьбу за существование, но во всех этих случаях мы имеем типичное переживание особей, наиболее приспособленных к данным условиям существования и, в частности, наиболее успешно сопротивляющихся указанным элиминирующим факторам.
Несмотря на различия в элиминирующих факторах и на различия в частных результатах, следует отметить, что борьба за существование в форме индивидуальной элиминации происходит всегда и везде, при всяких условиях. Даже в искусственно создаваемых постоянных оптимальных условиях внешней среды в лабораторных культурах никогда не прекращается действие естественного отбора, ибо борьба за существование определяется не одними лишь внешними факторами, а соотношениями между организмом и средой. При наличии же индивидуальных различий и индивидуальной изменчивости соотношение между организмом и средой является для разных особей различным. Элементарный отбор на жизнеспособность, связанный с устранением неблагоприятных мутаций, происходит всегда и при всех условиях, и это есть основное, наиболее общее выражение индивидуальной элиминации.
По элиминирующим факторам можно различать прямую элиминацию физическими или биологическими факторами и косвенную элиминацию физиологическими факторами.
1. Прямая элиминация физическими факторами внешней среды, или климатом в широком смысле, ведет к переживанию особей, наиболее защищенных от неблагоприятных влияний внешней среды, т. е. либо наиболее стойких, либо способных вовремя избегнуть этих влияний, либо обладающих специальными средствами защиты.
2. Прямая элиминация биологическими факторами, т. е. хищниками, паразитами и патогенными микроорганизмами, ведет к переживанию особей, наиболее вооруженных активными или пассивными средствами защиты против хищников и паразитов, а также к переживанию особей наиболее иммунных к болезням. Уничтожение биологическими факторами имеет огромное значение. Вместе с предыдущей формой элиминации она ведет к смене приспособлений (алломорфоз) соответственно изменениям среды, но также и к общему повышению организации (ароморфоз) или иногда и к общему повышению плодовитости и дегенерации (катаморфоз).
3. Косвенная элиминация физиологическими факторам и, и именно главным образом голодом, ведет обычно лишь косвенно через ослабление организма, к частичному или полному устранению от размножения (частью и в результате половой конкуренции). При ограниченности пищевых ресурсов для данного вида организмов численность последних ограничивается на данной территории известными пределами. Всякое приближение к этому пределу ведет к нехватке пищи для большинства особей. Ослабление организма вследствие голода (иногда только зимнего) ведет к ослаблению функций размножения и к повышенной смертности от неблагоприятных физических влияний, а также к усиленному истреблению хищниками, паразитами и болезнями. Интенсивность элиминации возрастает, следовательно, с увеличением плотности населения.
И в этом случае происходит отбор наиболее приспособленных особей, наиболее успешно добывающих себе пропитание и наиболее экономно его использующих. Это ведет у хищников к развитию органов чувств и средств нападения, а также соответствующих инстинктов.
Так как при расхождении форм и специализации по роду пищевого материала использование последнего делается более совершенным, а конкуренция между расходящимися формами снижается, то физиологическая элиминация ведет к прогрессивной специализации (алломорфоз и теломорфоз).
При борьбе, обусловленной нехваткой пищевого материала, приобретает очень большое значение истощение организма (особенно самки) при интенсивном образовании половых продуктов. Поэтому известные преимущества в борьбе за существование получают менее плодовитые особи, как менее истощенные. В особенности при развитии крупных яиц и при наличии живорождения и различных форм заботы о потомстве (кормление) борьба за жизненные средства приводит к естественному отбору менее истощенных, т. е. менее плодовитых особей, в смысле сокращения числа одновременно откладываемых яиц или рождаемых детенышей. Сокращение плодовитости (при возможном продлении жизни и размножении) является вторым, после специализации, типичным результатом косвенной элиминации при ограниченности средств питания.
в. При семейной элиминации происходят те же явления прямо го или косвенного устранения целых семей действием биотических или абиотических факторов. Примитивные формы семейного отбора возможны в силу того, что потомство одной пары является в отношении своих наследственных свойств более однородным, чем вся популяции в целом. Поэтому сюда относится в известной степени и отбор на плодовитость, характерный для общей элиминации. Совместная жизнь родителей с их потомством значительно увеличивает значение семейного отбора, так как в этом случае члены одной семьи подвергаются в значительной мере одним и тем же опасностям. Поэтому вопросы элиминации и переживания будут решаться нередко для всей семьи в целом. В этом случае будет происходить естественный отбор наиболее стойких, наиболее активных и защищенных семей, причем особое значение приобретают различные формы заботы о потомстве, связанные с развитием сложной системы инстинктов.
г. При групповой элиминации происходит устранение целых популяций. Это может быть обусловлено истреблением хищниками или паразитами, а также неблагоприятными климатическими условиями.
Резкое изменение жизненных условий, как, например, вторжение и размножение нового хищника, может быстро привести к полному вымиранию данной формы в данной местности. Еще большее значение имеет, однако, косвенное устранение от размножения в результате обессиления в борьбе за пищу и другие средства для жизни и размножения. Действие указанных факторов приводит через постепенное сокращение численности данной популяции к ее вытеснению, т. е. размножению других популяций, более приспособленных к условиям существования в данной местности.
Поскольку условия эти различны в разных местностях, популяция, вытесняемая в одной местности, может получить распространение ь другой.
Групповая элиминация ведет к распределению конкретных популяций (а также целых экотипов, подвидов, видов) по различным стациям и биотопам и через уничтожение отдельных популяций (экотипов, подвидов, видов) также к образованию более резких разрывов между таксономическими группировками.
Характер борьбы за существование определяется всегда конкретными соотношениями между организмом и средой, как они проявляются в данное время, в данном месте и при данных условиях. Разные формы элиминации связаны при этом с различными формами естественного отбора (как ее обратной стороной) и определяют, таким образом, различные направления эволюционного процесса.
Индивидуальная элиминация физическими или биологическими факторами ведет при разнообразных условиях среды к развитию широких приспособлений типа ароморфозов. При интенсивном истреблении она же ведет к увеличению плодовитости, а при случайном характере элиминации, кроме того, к катаморфозам. При более однородных жизненных условиях та же прямая индивидуальная элиминация ведет к более частным приспособлениям типа алломорфозов. При косвенной индивидуальной элиминации, связанной с ограниченностью жизненных средств, эволюция идет по пути прогрессивной специализации (теломорфоз), связанной с понижением плодовитости и уменьшением пластичности данной формы организмов (И. Шмальгаузен, 1939а).
Семейная элиминация ведет к установлению оптимальной плодовитости (большой при интенсивной прямой элиминации и малой при интенсивной косвенной) и к развитию различных способов защиты яиц и молоди и различных форм заботы о потомстве.
Групповая элиминация определяет размещение конкретных популяций по экологическим нишам — стациям и биотопам, а также таксономическое расчленение мира организмов.
Мы видим, что дарвиновское понятие борьбы за существование может быть охарактеризовано с двух различных сторон: как элиминация особей различными факторами среды и как соревнование особей в их борьбе за жизнь. Расчленение понятия борьбы за существование может быть проведено как с одной, так и с другой точки зрения.
Формы борьбы за существование характеризуются прежде всего как различные формы соревнования, связанные также с различными направлениями естественного отбора и определяющие через это характер эволюционного процесса. Индивидуальное соревнование определяет изменения структуры и функций организма, межсемейное соревнование — эволюцию форм размножения, а межгрупповое соревнование — распределение организмов и таксономическое их расхождение.
С другой стороны, различные формы элиминации прекрасно характеризуют очень многие формы борьбы за существование, связанные с различными направлешшми естественного отбора, определяющими те или иные пути эволюции организмов (общее повышение организации, приспособление к частным условиям среды, специализацию и т. п.), эволюцию форм размножения и заботы о потомстве, а также географическое и экологическое распределение организмов.
Расчленение понятия борьбы за существование на основе двух указанных принципов может быть проведено гораздо дальше. Вопрос о том, какому принципу дать предпочтение, зависит как от конкретных задач, стоящих перед исследователем, так и от особенностей материала.
В первом аспекте все внимание фиксируется на организме, его распространении и его таксономических соотношениях. При этом анализируются строение и функции орга-пизма в их зависимости от всего комплекса факторов (условий жизни), составляющих внешнюю среду в целом. Дается сравнительная оценка строения и эволюции близких организмов в различных конкретных условиях существования. Задача м о р ф о -лога-филогенетика связывается здесь с палеонтологическими и таксономическими исследованиями.
Во втором аспекте все внимание фиксируется на элиминирующих факторах и вместе с тем проводится анализ факторов внешней среды (условий жизни), определяющих переживание и размножение данного организма в целом. При этом дается сравнительная оценка всех условий существования в эволюции конкретного организма. Таким образом, задачи морфолога-физиолога связываются здесь с экологическими исследованиями.
5. ИНТЕНСИВНОСТЬ СОРЕВНОВАНИЯ И ЕГО ИЗБИРАТЕЛЬНОЕ ЗНАЧЕНИЕ
Мерилом интенсивности элиминации является плодовитость организма, отнесенная к единице времени (количество продуцируемых в течение года яиц или семян). О плодовитости современных животных и растений мы имеем в общем достаточно полное представление. Казалось бы, что большая плодовитость, а следовательно, интенсивная элиминация, должна обеспечивать высокую эффективность естественного отбора и вместе с тем быстрые темпы эволюции. Однако большая плодовитость низших организмов вообще и низших представителей почти любого крупного таксономического подразделения (типа, класса) по сравнению с высшими его представителями находится в резком противоречии с таким предположением. Интенсивная элиминация, при прочих равных условиях, конечно, обеспечивает большую эффективность естественного отбора, если эта элиминация имеет избирательный характер. Однако элиминация может иметь и неизбирательный характер (общая элиминация случайными факторами. И. Шмдльгаузен, 1939а), а ее избирательное значение может не находиться в прямой связи с ее интенсивностью. Можно сказать даже наоборот — особо интенсивная элиминация лишь редко имеет явно избирательный индивидуальный характер. С другой стороны, весьма эффективная, строго индивидуальная элиминация достигается иногда при ничтожной ее интенсивности.
Эффективность естественного отбора определяется конкретным положением организма в окружающей его среде, т. е. не только интенсивностью его элиминации, а следовательно — плодовитостью, скоростью созревания и смены поколений данного вида организмов, но и характером его элиминации (в особенности — в различных возрастах) и активностью самого организма в борьбе за жизнь и за сохранение своего потомства.
Особое значение в этом взаимодействии между организмом и окружающей его средой имеет, с одной стороны, активность и мощность агрессивных факторов среды (в первую очередь — других организмов, т. е. хищников и паразитов) и, с другой стороны, степень активности самого организма в добыче жизненных средств, в защите своей жизни и жизни своего потомства. Чем более активны истребляющие факторы среды, тем более общий характер принимает элиминация, теряющая перед лицом стихийных сил природы или мощного агрессора свой избирательный характер. Значение индивидуальных особенностей отступает в этом случае на задний план. Вперед выдвигается значение случайных столкновений беззащитного организма с истребляющими факторами подавляющей силы.
С другой стороны, чем менее мощны истребляющие факторы и чем выше активность организма, тем большее значение приобретают индивидуальные особенности отдельной особи. Если организм уже может бороться с неблагоприятными физическими влияниями или успешно избегать встречи с врагами или даже активно от них защищаться, то нередко малейшие индивидуальные различия в общей силе, в инстинктах или в специальных средствах защиты решают вопрос о жизни или смерти особи. Элиминация приобретает избирательный характер. Точно так же элиминация имеет строго избирательный характер и при косвенном истреблении особей в результате недоедания, связанного с общей нехваткой пищевых средств. В этом случае активность в добывании жизненных средств и экономное их расходование получают решающее значение.
Мы видим, что избирательный характер элиминации возрастает с увеличением активности организма и с общим упрочением его положения в окружающей внешней среде. Интенсивность же элиминации как раз в этом случае несомненно падает. Сказанное может быть иллюстрировано данными Уортингтона (Worthington E. В., 1940) относительно значения активного хищничества для дивергенции форм среди африканских рыб. В озерах Виктория, Ньяса и Эдуард среди рыб отсутствуют активные крупные хищники (имеются только мало активные двудышащие и сомы). Соответственно в оз. Виктория рыбы рода Haplochromys дали начало множеству эндемичных форм, начиная с рыб с маленькими ртами и маленькими волосовидными зубами, питающихся микроскопическим планктоном, и кончая рыбами с большими ртами и зубами, питающимися другими рыбами. Промежуточные виды питаются моллюсками и имеют нередко большие зубы с плоскими дробящими коронками. В оз. Эдуард множество форм образуют пелагические ципринодонты.
Очевидно, среди этих рыб преобладало значение активного соревнования, которое и привело к богатому экологическому расхождению. Элиминация имела здесь избирательный характер.
С другой сторны, в озерах Альберт и Рудольф имеются большие активные хищники: «нильский окунь» и «рыба тигр». Соответственно этому другие рыбы, которыми эти хищники кормятся, не дали заметной радиации форм. Очевидно, их прямая истребляемость имеет почти неизбирательный характер. Борьба за существование оказывается в значительной мере пассивной.
Отмечается также удивительный консерватизм двудышащей рыбы Protopterus, которая ведет весьма малоактивный образ жизни и в то же время не дала заметного видообразования. Другие примеры будут разобраны далее. Здесь мы должны только подчеркнуть, что избирательная элиминация, а следовательно и отбор, захватывают, конечно, организм в его морфофизиологическом выражении, т. е. по фенотипу. Непроявляющаяся мутация не может иметь избирательного значения. Поэтому все формы скрытого мутирования, связанного с рецессивностью мутации в гетерозиготном диплоидном, а тем более тетраплоидном организме, или с высокой регулярторной способностью организма, не являются непосредственным объектом отбора. Однако они могут иметь косвенное значение, определяя стойкость организма в известных условиях и его пластичность, при их изменении. Об этом будет речь в дальнейшем изложении.
Естественный отбор является процессом, обратным по отношению к элиминации, однако только по отношению к избирательной элиминации, да и то с известными оговорками. Поэтому острота элиминации ни в какой мере не определяет интенсивности естественного отбора. Это создает трудности для учета реальных интенсивностей отбора в природных условиях. Об этой интенсивности можно иметь суждение лишь по реальной скорости распространения известной формы в известных условиях существования. Но указанная скорость определяется и другими факторами, которые нам еще предстоит рассмотреть.
Математическая теория естественного отбора, разработанная Фишером (Fisher R. А., 1930), Холденом (Haldane J. S., 1924), Райтом (Wright S., 1931) и др., исходит из теоретических предпосылок и по необходимости крайне упрощает стоящую перед нею задачу. Однако и при таком упрощении задачи большинство вопросов требует для своего разрешения чрезвычайного напряжения математических средств. Мы можем в дальнейшем разборе принять во внимание лишь некоторые основные выводы этих авторов, касающиеся вопросов соотношения мутабильности, отбора и изоляции в процессах изменения генетической структуры популяций. Здесь же мы отметим лишь некоторые качественные характеристики элиминации и отбора, на которые пока обращалось недостаточно внимания, хотя они имеют большое значение.
Нередко говорят об уничтожающей роли естественного отбора вообще, или об отрицательном отборе в специальном случае уничтожения неприспособленных форм. Во всех случаях уничтожения отдельных особей или целых групп особей я предпочитаю говорить об элиминации. При элиминации оформленных таксономических групп (рас, подвидов, видов и высших таксономических подразделений) можно говорить о вымирании. Термин «отрицательный отбор» отличается двусмысленностью, так как в применении к известным признакам он получает противоположное значение: элиминация ветром летающих насекомых небольших океанических островов («отрицательный отбор») приводит к переживанию, т. е. положительному отбору нелетающих особей. В данном случае также нередко говорят, что редукция крыльев есть результат «отрицательного отбора» (по недоразвитым крыльям).
Во всех случаях уничтожения отдельных особей или местных популяций мы будем говорить об их элиминации.
Межгрупповое соревнование, связанное с элиминацией целых групп особей (популяций, подвидов, видов и др.), ведет к переживанию одних форм (отбор) и уничтожению других (вымирание). В этом случае отбор происходит между уже существующими формами. Он не имеет здесь явного творческого значения. Говорят об отсеивающей роли отбора (сравнение с решетом) или об его уничтожающей роли, так как в результате его действия число групп и, следовательно, многообразие органических форм уменьшается. Таково значение отбора при межгрупповом соревновании. Сюда относится дарвиновское вымирание промежуточных форм, ведущее к таксономическому расхождению организмов. Так как группа, уничтожаемая или вытесняемая в одной местности, может получить преобладание в другой, то межгрупповое соревнование (групповая элиминация) связано с распределяющей ролью отбора. Однако территориальное и экологическое распределение организмов связано с их приспособленностью к известным условиям существования. В этом случае мы имеем дело с положительным фактором эволюции флор и фаун, т. е. именно С деятельностью естественного отбора. Однако последний оперирует с готовыми формами и, следовательно, не имеет здесь творческого значения.
Определенно творческое значение имеет лишь индивидуальный естественный отбор, основанный на соревновании особей внутри известной группы (популяции, расы, подвида, вида), т. е. на индивидуальной элиминации. Он действует всегда на фоне подвижного материала, непрерывно меняющегося в процессах мутирования и комбинирования. Последнее определяется скрещиванием. Комбинирование и даже само мутирование идет под постоянным контролем естественного отбора. Благоприятные комбинации не возникают случайно — они создаются под направляющим действием естественного отбора. Именно эта роль естественного отбора, творящего новые формы, и составляет предмет нашего рассмотрения (в остальном см. И. Шмальгаузен, 19386, 1939а).
Творческая роль естественного отбора проявляется также в различных формах в зависимости от данных соотношений между организмом и факторами окружающей среды.
При изменении условий внешней среды приспособленность организма к этой среде, естественно, теряется, и его элиминация возрастает. При наличии таких индивидуальных уклонений, которые повышают приспособленность организмов к новым условиям существования, они будут переживать в большем числе и в результате размножения получать все большее распространение в популяции. Особи, представляющие прежнюю норму, окажутся в худшем положении и будут гибнуть в относительно большем числе. Популяция начнет изменяться приспособительно к новым условиям существования и таким образом установится новая норма на месте старой.
Это — обычная дарвиновская борьба за существование, связанная с прогрессивной эволюцией на основе элиминации прежней нормы. Естественный отбор выступает здесь в роли ведущего фактора эволюции, изменяющего старую и создающего новую, в данном случае более приспособленную, норму.
Если же факторы внешней среды получили довольно устойчивый характер и организм уже вполне приспособлен к данным условиям существования, то преимущества в борьбе за существование будут на стороне установившейся нормы, а уклонения, нарушающие приспособленность организма, начнут гибнуть в гораздо большем числе.
Несмотря на наличие дарвиновской борьбы за существование, заметных изменений в организации не произойдет. Наоборот, естественный отбор будет поддерживать существование данной нормальной организации (хотя и будут происходить изменения в процессах индивидуального развития, приводящие к увеличению его устойчивости). Отбор выступает здесь в роли стабилизирующего фактора, действующего на основании э л и м и н а-ц и и всех уклонений от нормы.
Таким образом, две формы элиминации определяют два различных выражения естественного отбора — его ведущую роль (меняющую норму) и его стабилизирующее значение (укрепляющее установившуюся норму). Если бы не указанная двусмысленность в понимании «отрицательного» отбора — 1) как элиминации менее приспособленных особей вообще и 2) как отбора на уменьшение известных признаков, то, принимая второе определение, можно было бы различать следующие основные формы отбора:
I. Отбор положительный (ведущая форма), идущий в направлении нарастания и усложнения признака. В результате происходит прогрессивное развитие данной части.
П. Отбор отрицательный (ведущая форма), идущий в сторону уменьшения и упрощения признака. В результате происходит регрессивное развитие данной части.
III. Отбор нейтральный (стабилизирующая форма), поддерживающий установившуюся нормальную величину и строение признака. В результате происходит стабилизация формообразования, т. е. автономизация онтогенеза и развитие его регулятор-ных механизмов.
6. ФОРМЫ ОТБОРА
Естественный отбор меняет не только свое направление, но и свои формы в зависимости от изменений условий и форм борьбы за существование. В борьбе за существование находят свое многообразное выражение сложнейшие взаимодействия между организмом и окружающей его неорганической и органической средой (включая и взаимозависимости между особями одного и того же вида организмов). Поэтому как борьба за существование, так и естественный отбор изменяются в своей интенсивности и в своей качественной характеристике при изменении любых факторов внешней среды, так же как и при изменении строения, функций и нормы реакций самого организма.
Мы здесь не ставим своей задачей анализ всех возможных соотношений, но фиксируем наше внимание на значении характера самих изменений (неуклонных, т. е. направленных, периодических или колеблющихся) однородной или разнородной среды, а также на изменчивости лабильных и стабильных организмов, обнаруживающих в своем формообразовании различную степень зависимости от факторов внешней среды.
Ведущая (движущая) форма отбора реализуется на основе селекционного преимущества (в измененных условиях внешней среды) определенных уклонений организации перед нормой, установившейся в прежних условиях существования. Она связана с частичной элиминацией прежней нормы и установлением новой. Стабилизирующая форма отбора осуществляется на основе селекционного преимущества (в известных, особенно в колеблющихся, условиях) нормальной организации перед уклонениями от нормы. Она связана с элиминацией большинства уклонений и с выработкой более устойчивых механизмов нормального формообразования.
А. ВЕДУЩАЯ ФОРМА ОТБОРА
а. Стабильный организм в однородной неуклонно меняющейся среде. Наиболее простыми являются изменения соотношений между однородной средой, меняющейся в одном определенном направлении, и изменчивым организмом со стабильным (т. е. независимым от изменений внешних факторов) формообразованием. Это — классическая форма естественного отбора, к которой относится и большинство его доказательств. Его действие можно пояснить следующими схемами (рис. 9, А, В).
Первый рисунок (А) показывает типичную вариационную кривую (1), характеризующую известный признак всех особей некоторой популяции определенного вида организмов. Вторая кривая (2) характеризует изменчивость того же признака «нормальных» особей, переживающих и оставляющих потомство. Промежуток между обеими вариационными кривыми представляет зону вариаций, элиминируемых в борьбе за существование.

Рис. 9. Отбор в однородных условиях внешней среды. А — отбор при установившихся условиях (стабилизирующая форма отбора); 1 — вариационная кривая всех особей; 2 — вариационная кривая особей, выживающих и оставляющих потомство («норма»). Заштрихована область вариантов, элиминируемых при данных условиях. В —отбор при изменении этих условий в определенном направлении (движущая форма отбора): 1 — вариационная кривая всех особей; 2 — вариационная кривая особей, выживающих и оставляющих потомство в новых условиях (новая «норма»); С, D — другой план изображения механизма действия стабилизирующей и движущей форм отбора
При изменении внешней среды в определенном направлении, нарушающем приспособленность организма по данному признаку, некоторые особи прежней нормы окажутся менее приспособленными, а некоторые варианты, ранее устранявшиеся, могут приобрести известные преимущества. В результате этого, с одной стороны вариационной кривой (рис. В) произойдет сокращение численности переживающей нормы и соответственное расширение зоны элиминируемых вариаций ( —), а с другой, благоприят-ствуемой, стороны произойдет расширение нормы за счет зоны вариаций, которые ранее элиминировались ( + ). Кривая переживающих особей примет асимметричный вид и средняя сдвинется в благоприятствуемую сторону (мода может вначале оставаться на месте и в своем движении отстает от средней тем более, чем резче меняется среда и чем жестче элиминация). Накопление вариаций, а следовательно — и мутаций, которые ранее элиминировались, ведет к некоторому нарушению регуляторного аппарата («генного баланса» и всей системы морфогенетических корреляций вообще), а это, в свою очередь, способствует проявлению мутаций, которые были погашены в своем выражении (вскрытие резервов, см. Н-5), и дальнейшему повышению мутабильности. Поэтому вариационная кривая расширяется, и тогда начинается ее движение в благоприятствуемом направлении за пределы вариантов, наблюдавшихся ранее (рис. У). Этот процесс идет медленнее, чем первая фаза эволюции,—сдвиг средней за счет наличных уже вариантов и мобилизации скрытых резервов внутривидовой изменчивости. Однако в благоприятных условиях эволюция приобретает затем характер самоускоряющегося движения и может идти все же с очень значительной скоростью (см. раздел IV). То же самое можно иллюстрировать и диаграммами, представляющими как бы поперечное сечение трехмерных вариационных кривых (рис. 9, С, D).
Большинство прямых и косвенных доказательств существования естественного отбора касается именно этой наиболее простой и наглядной его формы. Из косвенных доказательств можно сослаться на явления покровительственной окраски и мимикрии, а также на другие средства пассивной защиты. Совершенно ясным примером того же действия естественного отбора может служить редукция крыльев у островных насекомых или редукция органов зрения у роющих позвоночных.
Из прямых доказательств ведущей роли естественного отбора у стабильных организмов можно упомянуть наблюдения Уэлдона (Weldon, 1898) над крабами Carcinus maenas в гавани Плимута, где, в связи с постройкой мола и замутнением воды, произошло сужение головогрудного щита (очевидно, это лишь первая фаза процесса — сдвиг средней в пределах имевшихся уже вариаций). Сюда же относятся наблюдения Гаррисона (Harrison, 1920) над изменением окраски бабочек Oporabia autumnata в сторону посвет-ления при замене соснового леса березовым. Наконец, из новейших исследований такого рода следует упомянуть наблюдения Дубовского (1941) над приспособлениями пресноводных и морских ракушковых рачков (Ostracoda), сопровождавшиеся экспериментальными исследованиями значения некоторых адаптации (средств прикрепления) и отбора в условиях текучей воды.
б. Лабильные организмы в однородной неуклонно меняющейся среде. В случае эволюции лабильного организма наша простая схема полностью сохраняет свое значение, но усложняется лишь за счет наличия модификационных изменений, прикрывающих мутационные изменения и потому затрудняющих их учет. Ведущая форма отбора комбинируется здесь со стабилизирующей, о которой речь будет идти в дальнейшем. Все же направленность эволюции бывает ясно выражена, и движущая роль отбора не вызывает сомнений. Зона элиминируемых вариаций составляется в этом случае, кроме мутаций, также из модификаций (из морфозов и адаптивных реакций на случайные, кратковременные изменения в факторах среды, т. е. из реакций, оказавшихся «неудачными» или «ошибочными»). Отбор положительных наследственных уклонений происходит на фоне более или менее значительных и более или менее совершенных индивидуальных приспособлений (модификаций).
Многие географические изменения, отличающиеся своей непрерывностью, характеризуются именно теснейшим соединением мутационных и модификационных изменений, идущих в одном и том же направлении.
Так, например, чистые линии льна, высеянные в разных широтах, дают фенотипические изменения, сходные с географическими формами данных мест, отличающиеся, однако, и по своему генотипу. Северный лен-долгунец отличается большей высотой и меньшим числом стеблей и коробочек, чем южный лен-кудряш. Это — наследственные отличия. Однако на юге лен-долгунец становится ниже, а число стеблей и коробочек увеличивается. Наоборот, на севере лен-кудряш до известной степени приближается к долгунцу. Сон-прострел (Anemone Pulsatilla) обладает в своей западной расе разбросанными и мелко рассеченными листьями, а также поникшими цветами. У восточной расы листья приподняты, более грубо рассечены и цветы стоячие. Это — наследственные различия. Их гибриды менделируют. В их потомстве происходит расщепление по форме листьев в отношении 25% особей, обладающих листьями с широкой верхушкой, 25 % — с узкой и 50 % — со средней верхушкой, как у гибридов. Однако эти наследственные различия перекрываются сходными модификационными изменениями, которые могут быть получены в опыте при изменении интенсивности освещения (соответственно меньшей облачности восточной Европы) !.
Северные млекопитающие и птицы отличаются более крупными размерами от южных представителей той же группы (правило Бергманна). Эти отличия — наследственные, и они могли установиться в результате естественного отбора на более экономичный обмен и меньшую потерю тепла у северных форм. Однако частично эти различия могут иметь характер модификаций, как показывают некоторые эксперименты. Точно так же более короткие уши и хвосты северных грызунов и некоторых других млекопитающих (правило Аллена) являются наследственными свойствами, развившимися в процессе естественного отбора на меньшую потерю тепла (через уши и хвост, обладающие слабым шерстяным покровом и служащие для теплорегуляции). То же самое касается густоты общего шерстяного покрова, длины волос и их толщины у северных млекопитающих по сравнению с южными расами и подвидами. Однако и здесь наследственные изменения перекрываются сходными модификационными (как видно из опытов Семнера).
в. Стабильные организмы в разнообразной неуклонно меняющейся среде. Естественный отбор в однородной среде, конечно, никогда не осуществляется в чистом виде. Наиболее единообразны разве лишь условия существования паразитов, частично — глубоководных или пещерных животных, животных, зарывающихся в более глубокие слои почвы или в грунт более глубоких вод.
За этими лишь исключениями, факторы среды меняются в зависимости от места пребывания, от времени года и даже суточных или случайных колебаний освещения, температуры, влажности и т. п. Внешняя среда не бывает однородной ни в пространстве, ни во времени, ни по своим физическим факторам, ни по биологическим (пища, враги, паразиты).
В этом случае наша схема ведущего отбора усложняется в том отношении, что никакая «норма» не может быть одинаково приспособленной и ко всем местным условиям среды и ко всем ее как случайным, так и периодическим изменениям.
Поэтому само понятие «нормы» становится несколько неопределенным. Мы все же будем понимать под «нормой» просто всю сумму особей, которые фактически переживают в конкретных (разнообразных) условиях существования и оставляют после себя потомство. Это, однако, лишь условная средняя норма, которая фактически непрерывно меняется.
Кроме зоны безусловно элиминируемых уклонений, теперь приходится учитывать существование зоны условно благоприятных уклонений, которые в некоторых частных условиях существования получают известные преимущества перед средней нормой. В некоторых местностях, в известные сезоны или при случайных уклонениях в факторах среды переживают и оставляют потомство не представители средней нормы, а эти условно благоприятные варианты. В других местностях, при других уклонениях эти варианты элиминируются, но зато переживают другие варианты. При дружной весне все преимущества будут на стороне рано и быстро развивающихся растений и рано вылупляющихся личинок насекомых (обладающих низким температурным порогом реактивности). Однако в следующий год вернувшиеся весной морозы могут убить их потомство, и тогда все преимущества будут на стороне поздно и медленно развивающихся особей. Отбор меняет свое направление, и это поддерживает весьма значительную гетерогенность популяций растений и животных, живущих в разнообразной и непостоянной среде.
В случае существования ясно очерченных локальных (экологических) условий гетерогенность популяций принимает формы более или менее ярко выраженного генетического полиморфизма (гетероморфизма). Так должен был развиться гетероморфизм богомолов (Mantis religiosa), обладающих зелеными и буро-желтыми формами, которые прекрасно защищены в зеленой листве, на бурых стеблях кустарников и на желтой, высыхающей летом траве. Эти основные формы имеют наследственный характер и непрерывно выщепляются в чистом виде у родителей любой окраски (М. М. Беляев, 1946).

Рис. 10. Основные типы окраски у божьей коровки Harmonia axyriiis. а — var succinea, b— var frigida, с — var 19-signata, d — var aulica, e — var axyridis, f — var spectabilis, g — var conspicua. По Ф. Добжанскому, 1937
Таково же значение гетероморфизма мимикрирующих самок африканских папилионид (Papilio polytes и P. dardanus), которые находят свою защиту в сообществе различных иммунных видов (относящихся к родам Amauris и Danais). Каждая форма имеет свою самостоятельную линию эволюции, как это показывает факт образования у P. dardanus в Ю. Африке ряда локальных географических форм, копирующих местные подвиды Amauris echeria. Между тем это один вид, непрерывно выщепляющий различные, характерные для данной местности, формы самок.
Холден указывает, что в некоторых случаях гетероморфизм может быть очень устойчивым. Так, в современных популяциях садовых улиток Cepea hortensis и С. nemoralis встречаются гладкие (однотонные) и полосатые формы. Последние несколько преобладают. В отложениях неолита типы этих же улиток найдены почти в современных пропорциях. В плейстоцене оба типа были приблизительно равночисленными. Следовательно, если и имеется определенный сдвиг, он совершался очень медленно (Lang, 1912).
В других случаях нетрудно установить не только географические, но и сезонные изменения направления отбора. У божьих коровок имеется множество вариантов, выражающихся в окраске (рис. 10). Некоторые виды оказываются резко гетероморфными. Так, Ф. Добжанский изучал географические расы Harmonia axyridis, отличающиеся необыкновенным разнообразием в положенной распространении пятен. Почти во всех расах повторяются одни\ и те же красочные типы, однако в различных пропорциях. При расселении этого вида в одних местностях получали условное преимущество одни формы, в других — другие. Тимофееву-Ресовскому удалось показать даже изменение направления естественного отбора по сезонам. У божьей коровки Adalia bipunctata в осенней популяции преобладает «черная» форма. Однако, очевидно в силу большей ее смертности во время зимовки, весной преобладает «красная» форма. По Лукину (1939), у клопа Pyrrhocoris apterus наблюдаются и локальная и сезонная изменчивость размеров тела.
г. Лабильные организмы в разнообразной неуклонно меняющейся среде. При изменении лабильного организма в разнообразной среде картина эволюции усложняется наложением модификации на существующий уже генетический полиморфизм.
Географические изменения высших растений происходят главным образом по этому типу комбинирования естественного отбора в гетероморфных популяциях (с обособлением «экотипов») и прямого приспособления (выражаемого в «экофенах», по терминологии Турессона). Однако эволюция птиц и млекопитающих также идет в значительной мере по этому пути.
Чем выше значение адаптивных модификаций, тем менее заметную роль играет (на данном отрезке времени) естественный отбор. Руководящая роль переходит тогда на время к адаптивной модификации.
Б. СТАБИЛИЗИРУЮЩАЯ ФОРМА ОТБОРА
Стабилизирующая форма отбора ведет, через элиминацию всех случайных уклонений, к повышению устойчивости существующей уже или устанавливающейся нормы. Б этом случае имеется в виду, что все организмы до известной степени лабильны, т. е. так или иначе, по меньшей мере на ранних стадиях развития, реагируют на изменения в факторах внешней среды, причем эти реакции далеко не всегда оказываются благоприятными (особенно если эти реакции были ответом на случайные и кратковременные изменения). При изменении факторов внешней среды, а также в процессе мутирования, устойчивость формообразовательных систем нарушается (они лабилизируются). В процессе эволюции они в той или иной мере восстанавливаются благодаря непрерывной деятельности стабилизирующего отбора. Результатом этой деятельности является стабилизация форм вследствие создания регу-ляторного аппарата, защищающего нормальное формообразование от возможных нарушений со стороны случайных уклонений в факторах внешней среды, а также и со стороны небольших уклонений во внутренних факторах (т. е. мутаций). Естественные отбор идет в этом случае на основе селекционного преимущества нормы (в частности — новой нормы) перед любыми от нее уклонениями.
а. Стабилизирующий отбор в однородных условиях. В природе лишь редко встречаются виды организмов, живущие во вполне однородных условиях внешней среды. Это касается в известной степени лишь специализированных паразитов, глубоководных животных и других подобных организмов, живущих в весьма ограниченной среде.
Поэтому мы ограничимся вначале лишь общим рассмотрением механизма стабилизирующего отбора в этих наиболее простых условиях, реально всегда осложняемых существованием локальных или сезонных (б), а также случайных (в) изменений в факторах внешней среды. Известные наблюдения Бумпуса над воробьями, погибшими в результате бури (Bumpus, 1899), ясно показывают наличие стабилизирующего отбора в природе. Биометрические исследования погибших воробьев показали, что большинство из них представляло в том или ином отношении уклонения от обычной нормы. Ясно, что именно нормальная организация оказалась наиболее устойчивой при несколько большем напряжении жизненных сил, которого потребовала внезапно налетевшая буря.
Роль стабилизирующего отбора, охраняющего норму, видна в особенности ясно при более детальном генетическом анализе процессов, постоянно протекающих в хорошо изученном материале опытных культур дрозофилы (Drosophila melanogaster), а также при анализе генетического состава природных популяций той же дрозофилы (то же доказано и для других ее видов).
Процесс мутирования, приводящий обычно к более или менее заметному уклонению от «нормального» фенотипа, находит противодействие в естественной элиминации всех вредных уклонений. Однако малые уклонения, лежащие еще в пределах условной нормы (в гетерозиготном состоянии), т. е. фактически способные созреть и оставить потомство, полностью не устраняются и участвуют через скрещивание в постоянной перестройке генотипа данной популяции. Если вредные (условно вредные) уклонения прямо не элиминируются, то они постоянно обезвреживаются через отбор наиболее жизненных комбинаций. Это показывают многочисленные опыты культивирования различных мутаций дрозофилы в лаборатории, при которых степень выражения вредной мутации непрерывно падала. Что это было результатом отбора (т. е. преимущественного переживания и размножения особей, менее уклоняющихся от нормы), видно на опыте скрещивания разных линий с угасшим выражением этой мутации (например, eyeless или stubbloid). После такого скрещивания выражение мутации по меньшей мере частично восстанавливалось. Это означает, ч\го небольшие наследственные уклонения, подавляющие выражение культивируемой мутации («гены-модификаторы») и отобранные во время ведения изолированных культур обеих линий, при скрещивании большей частью перешли в гетерозиготное состояние (так как в различных линиях в большинстве возникали и отбирались разные модификаторы). В гетерозиготе модификаторы потеряли свое выражение и основная мутация вновь проявлялась. Такие же факты были установлены и при культивировании мутаций бокоплавов (Gammarus chevreuxi).
При устранении специфического выражения вредной мутации мы имеем восстановление основных выражений «нормального» фенотипа (при измененном генотипе). Это восстановление достигнуто систематической элиминацией всех более резких уклонений от нормы, т. е. стабилизирующим отбором особей наиболее жизнеспособных, или, иными словами, наиболее приближающихся к установившейся норме.
Существование того же самого процесса установлено, однако, и в популяциях дрозофилы. Природные популяции оказываются насыщенными огромным количеством мутаций (Четвериков, 1926; Тимофеев-Ресовский, 1927; Р. Берг и др., 1941, 1944а, б. См. также сводку Н. Дубинин, 1940а). Кроме рецессивных мутаций, в популяциях оказываются и многие доминантные мутации, близкие к нейтральным или даже вредные (Р. Берг), выражение которых подавлено в генотипе данной популяции или значительно снижено их проявление (Гершензон). Популяция сохраняет свой нормальный «дикий» фенотип, несмотря на непрерывное мутирование, сопровождающееся именно накоплением мутации и, следовательно, непрерывной перестройкой ее генотипа.
Это и есть механизм стабилизирующего отбора, непрерывно «охраняющего» норму и восстанавливающего ее при нарушениях, вызываемых отдельными, не слишком вредными или условно «нейтральными» (в гетерозиготном состоянии) мутациями. При погашении выражения мутации несомненно возрастает и индивидуальная стабильность, которая у «нормальной» особи всегда выше, чем у мутанта.
Если все не слишком вредные, т. е. условно «нейтральные», мутации в процессе дальнейшего мутирования и отбора малых мутаций («модификаторов») теряют свое выражение (в гетерозиготе), т. е. становятся вполне рецессивными, то фенотип не меняется, несмотря на наличие мутаций. Это означает развитие рецессивности мутации и доминантности нормы (в процессе эволюции) и является результатом стабилизирующего отбора.
б. Стабилизирующий отбор в разнородных локально или сезонно меняющихся условиях. Эволюция организмов, живущих в меняющихся условиях разнообразной внешней среды, не Ограничивается выработкой одной ограниченной нормы. В наиболее часто встречающихся типичных условиях преимущества в борьбе за существование будут на стороне некоторой средней нормы, Однако в других реально встречающихся условиях преимущества будут на стороне тех или иных уклонений от этой главной нормы. Точно так же и в различные сезоны требования, предъявляемые к организму, закономерно меняются. Изменяются и направления естественного отбора, и это приводит к установлению «второстепенных норм», т. е. известного полиморфизма (экологического или сезонного), который может иметь генетический характер (гетероморфизм), но может быть и модификационным (собственно полиморфизм). Практически чаще всего происходит комбинирование известного гетероморфизма с выработкой наиболее адекватных форм реагирования. Некоторая зависимость процессов индивидуального развития от изменений во внешних факторах, т. е. известная лабильность, означает и неустойчивость и вместе с тем ненадежность формообразования.
В процессе эволюции наиболее существенные «адаптивные нормы» в известной мере стабилизируются. Это связано с развитием регуляторных механизмов, защищающих эти целостные приспособительные реакции от возможных нарушений со стороны случайных внешних влияний. И это можно рассматривать лишь как результат элиминации всех уклонений от установившихся «адаптивных норм», т. е. типичных форм приспособления к реально встречающимся частным условиям существования. Зависимые процессы развития принимают при этом авторегуляторный характер.
Таким образом, развитие «теневых» и «световых» форм листьев иногда индуцируется заранее внешним фактором. У некоторых деревьев оно определяется условиями освещения в почках, заложившихся в прошлом сезоне вегетации (Nordhausen, цит. по Е. Ungerer, 1926), а затем полностью регулируется внутренним механизмом развития. В этом можно видеть защиту приспособительного формообразования организма от «ошибочных» реакций на переменчивые условия весеннего освещения, т. е. известные черты автономизации развития, которые объясняются только естественным отбором результатов «удачных» реакций, или, иначе говоря, элиминацией всех неблагоприятных уклонений (т. е. «ошибочных» модификаций).
У амфибиотических растений вырабатываются две-три адаптивные нормы для жизни в воде, на болоте или на суше. Они имеют вполне целостный характер и осуществляются при посредстве внутреннего авторегуляторного механизма развития, хотя частично или сполна детерминируются известными интенсивностями внешних факторов. При этом водные листья индуцируются вовсе не обязательно водной средой. У водяной гречихи (Polygonum amphibium) можно получить типичные «плавающие» листья во влажном воздухе, а не только в воде. У стрелолиста (Sagittaria sagittifolia) типичные лентовидные водяные листья развиваются и в воздухе при ослабленном освещении (Vischer, цит. по К. Goebel, 1928). Таким образом, детерминирующим фактором развития здесь является не тот фактор среды, который вызвал к жизни данную модификацию. В процессе эволюции выработался новый механизм индивидуального развития, что, очевидно, возможно только через естественный отбор. Так как отбор происходил на фоне адаптивной модификации, то он, во всяком случае частично, проявлялся в виде стабилизирующего отбора. В данных случаях известная степень влажности и затененности была, очевидно, более надежным раздражителем, чем переменчивый уровень воды в заболоченной местности. Смена детерминирующих факторов должна была произойти в результате элиминации всех неудачных реакций (т. е. «ошибочных» адаптации, приводивших к развитию водной формы на суше, или наоборот).
Подобная стабилизация адаптивных норм наблюдается и у животных, однако, главным образом, в качестве сезонных форм (подвижность большинства животных не ставит перед ними вопроса о локальных экологических адаптациях). Вполне стабильными являются сезонные формы бабочки Araschnia levana-prorsa. Довольно стабильны также, и во всяком случае вполне характерны для различных видов, и сезонные формы коловраток и дафний. Все они определяются в своей реализации не столько внешними факторами, сколько существованием определенных систем внутренних регуляторных механизмов. Авторегуляторный характер развития сезонных форм совершенно ясен. Нужно думать, что и здесь регуляторные механизмы развились в результате действия стабилизирующего отбора при устранении всех уклонений от приспособленной в данных условиях частной (сезонной) нормы.
в. Стабилизирующий отбор в колеблющихся условиях внешней среды. В любых условиях существования внешние факторы подвержены более или менее значительным случайным и кратковременным изменениям (нерегулярным колебаниям интенсивности). Однако в тех случаях, когда эти колебания очень резко выражены, стабилизация формообразования приобретает исключительно большое значение. Лабильные организмы реагируют на эти изменения, однако новые изменения в факторах среды лишают эти реакции их приспособительного значения. Реакция оказывается «преждевременной» и потому «ошибочной». У лабидьных организмов подавляющая масса неблагоприятных уклонений составляется из таких «ошибочных» модификаций, и потому интенсивная их элиминация должна вести к максимальной стабилизации форм. Стабилизирующий отбор должен идти с большей скоростью.
Возможность преобразования лабильного признака в стабильный показана экспериментально М. Камшиловым путем отбора на максимальное выражение при противоположных изменениях условий развития. Искусственным отбором удалось достигнуть того, что мутация eyeless выражалась одинаково как при воспитании на влажном корме (нормальный максимум), так и при воспитании на сухом корме (нормальный минимум). Получена экспериментальная автономизация развития данного признака, превратившегося из зависимого в независимый от изменений в факторах внешней среды (М. Камшилов, 1939в).
В природных условиях наиболее значительны колебания температуры, влажности и инсоляции в горных местностях, в резко континентальных и, наконец, в полярных странах.
В этих условиях действительно происходит иногда весьма быстрая стабилизация новых форм.
Н. И. Калабухов и В. М. Родионов (1936) показали, что равнинные предкавказские лесные мыши при перенесении в горы дают типичную горную реакцию в виде увеличения числа эритроцитов и количества гемоглобина в крови. Однако горные формы того же подвида Apodemus sylvaticus ciscaucasicus при перенесении на равнину уже не дают обратной реакции. Повышенное содержание гемоглобина у них наследственно стабилизировалось.
Обыкновенная пастушья сумка (Capsella bursa pastoris) имеет в горах Эрдшиас-даг в Малой Азии типичный альпийский характер — глубокие корни, низкий стебель, ксерофитный габитус с опушением. Хотя это растение, по Цедербауеру (Zederbauer, 1908, цит. по Weidenreich, 1921), могло быть занесено сюда только человеком и, следовательно, лишь недавно стало альпийским растением, оно при высеве семян в низменности в течение более четырех поколений все же сохраняло низкий стебель (корни и листья теряли альпийский характер). Следовательно, процессы формирования стебля здесь уже полностью стабилизировались (автономизировались).
Таких фактов частичного наследственного «фиксирования» изменений, которые ранее были зависимыми, т. е. имели характер приспособительных модификаций, известно очень много. Таковы различные экологические и географические формы как растений, так и животных, которые при переносе в иные условия, близкие к исходным, дают неполный возврат к прежнему фенотипу. Возможность возврата указывает на значение модификации в возникновении данной формы. То обстоятельство, что этот возврат является лишь частичным (или в известных признаках), показывает, что данный фенотип был все же объектом естественного отбора (и именно стабилизирующего, так как он происходил в пределах установившейся уже модифицированной нормы). Длинная и тонкая шерсть полярных млекопитающих (также, короткие уши и хвосты) есть наследственное их свойство. Однако при выращивании в более теплом климате эти признаки частично изменяются. С другой стороны, и млекопитающие умеренных стран получают в более холодном климате более длинную шерсть. Это — модификации, на фоне которых произошла стабилизация процессов развития более мощного шерстного покрова полярных животных (путем элиминации всех особей, которые в результате реакции на случайно более высокую температуру среды развивали недостаточный шерстный покров).
Если альпийский габитус развивается в горах у очень многих растений, то большинство типичных альпийских растений сохраняет хотя бы частично свой альпийский габитус и при переносе в долины. Формообразование их стало более стабильным, очевидно, в результате жесткой элиминации морозами и засухой всех особей, которые в случайно мягкие и влажные весны давали «ошибочную» долинную модификацию (с длинными стеблями, не-опушенными листьями и короткими корнями). Частным и притом более простым случаем стабилизации в условиях резких колебаний в факторах внешней среды является также распространение полиплоидных растений в горных, резко континентальных и полярных странах (Е. В. Вульф, 1937). Полиплоиды отличаются именно более устойчивым формообразованием.
В. СТАБИЛИЗАЦИЯ НОРМЫ В ПРОЦЕССЕ ЭВОЛЮЦИИ
Если мы рассматривали отдельно ведущую и стабилизирующую формы отбора, то это было сделано лишь для внесения максимальной ясности. На самом деле обе формы естественного отбора действуют всегда совместно, так как, с одной стороны, внешняя среда постепенно, но неуклонно меняется, и, следовательно, могут быть обнаружены ведущая роль отбора и наследственное изменение нормы, но, с другой стороны, это — процесс медленный и стабилизирующий отбор всегда и непрерывно ведет к развитию регуляторных механизмов, охраняющих эту медленно меняющуюся норму от нарушающих ее внешних влияний.
Ведущая форма отбора яснее всего устанавливается у стабильных организмов с автономным развитием, так как наследственные изменения у них не осложняются результатами индивидуального приспособления. Стабилизирующая форма отбора устанавливается и у «стабильных» организмов, но протекает более типично у лабильных организмов и особенно в условиях колеблющейся среды (с изменениями случайного характера). Однако именно в этих случаях трудно разграничить результаты ведущей формы отбора, меняющей норму реакций, и индивидуального приспособления, основанного на этих реакциях и дополняемого стабилизирующим отбором. Эта трудность нашла свое отражение и в нашем изложении: фактический материал, использованный в качестве иллюстраций, в сущности один и тот же. В некоторых случаях особенно ясно видно соединение обеих форм отбора в процессе эволюции (т. е. в процессе изменения самой нормы).
Известно, что при засухе листья вянут и затем опадают. В тропических странах нередко наблюдается вполне закономерный листопад в засушливое время года. Этот листопад является приспособлением, предохраняющим растение от потери воды при транспирации через листья и невозможности пополнения ее запаса через корневую систему. В умеренном климате зимой, при замерзании воды, создаются также условия «засухи», и это ведет также к листопаду (G. Nichols, 1924).
При экспериментальном удалении листьев у многих растений (Vicia faba, Sarothamnus scoparius, Lathyrus odoratus, Syringa vulgaris, Rosa centifolia, Robinia pseudoacacia и др.) сильно увеличивается число хлорофилльных зерен в ассимиляционных тканях стеблей и черешках листьев, а также происходит и новообразование хлоропластов в нижележащих слоях клеток. Клетки ассимиляционной ткани удлиняются в радиальном направлении, и возникает палисадная ткань (если ее не было; а если она была, то она сильнее разрастается). Число устьиц значительно возрастает. Экспериментально установлено усиление ассимиляции и транспи-рации (A. Boirivant, 1897, и Braun, 1899; цит. по Ungerer, 1926). Такая компенсация имеет, следовательно, приспособительный характер и приобретает особое значение у растений засушливых местностей, нередко сбрасывающих листья. В этом случае черешки и стебли функционально замещают сброшенные листья.
Сбрасывание листьев при засухе может оставлять растение безлистным на длительный период вегетации. Ксерофитное растение может в молодом возрасте развивать листья, а затем их окончательно терять (многие акации). Некоторые ксерофитные кустарники напоминают тогда безлистную метлу. Таковы некоторые спаржевые, но в особенности многие санталовые и мотыльковые, как, например, ракитник (Cytisus), бобровник (Spartium). Ассимилирующая ткань под кожицей стеблей обладает сильно редуцированными межклетниками. Устьица многочисленны, но малы и погружены. Здесь адаптивная модификация приобрела постоянный характер. В дальнейшем процессе эволюции побеги частью расширяются и образуют листообразные ветки — «филло-кладии» (например, иглицы — Ruscus hypoglossum и Ruscus acw leatus). У других растений расширяются не стебли, а черешки сбрасываемых листьев — «филл одни» (особенно у акаций). У Acacia heterophylla и др. в нормальном онтогенезе развиваются филлодии, и после этого опадают листья (рис. 11). Сбрасывание листьев и развитие филлодиев определяются уже автономно, т. е. внутренними факторами развития, а не засухой и компенсационной модификацией, связанной с отсутствием листьев. Таким образом, в процессе эволюции создается трудно понимаемая замена листьев другими сходными образованиями, несущими ту же функцию. Казалось бы, как это и происходит у других растений, сами листья могли бы приспособиться к условиям засухи.

Рис. 11. Молодое растение Acacia pyienantha. Первичные перистосложные листья (1–4) сменяются переходными листьями (5, 6) и развивающимися позднее типичными филлодиями (7 — 9). По К. Циммерман, 1930
В данном случае этот своеобразный путь эволюции объясним только как результат обусловленного засухой периодического листопада с последующими компенсационными явлениями. Здесь именно адаптивная модификация тканей стеблей и черешков имела ведущее значение в процессе дальнейшей эволюции. Эта модификация приобрела в ксерофитных условиях (очевидно, через механизм стабилизирующего отбора) значение постоянного, т. е. стабильного признака организации, развивающегося и без засухи. В процессе обычного отбора произошло затем приобретение листообразной формы новых органов, более приспособленных к ограниченной транспирации в сухой местности. Эти данные показывают с достаточной убедительностью, что именно вынужденная (периодической засухой и листопадом) адаптивная модификация была переходным этапом на пути эволюции, приведшей к образованию новых ассимилирующих органов (филлокладиев и филлодиев).
Стабилизация происходила, очевидно, на базе элиминации тех особей, которые давали в условиях случайно более влажной весны прежнюю реакцию нормального листообразования и затем, летом, страдали от чрезмерного испарения, пока не опадут листья, и от недостаточной ассимиляции углерода, пока не наступят компенсаторные модификации в стеблях и черешках. Здесь ясна и роль приспособительной модификации в процессе эволюции, и роль стабилизирующей формы отбора, ведущей к установлению внутреннего механизма развития этой модификации, и движущая роль обычного отбора, ведущего к развитию новой нормы с новыми листообразными органами.
Мощное развитие жевательной и височной мышц хищных млекопитающих сопровождалось прогрессивным развитием зубной системы, челюстей, венечного отростка нижней челюсти, скуловой дуги, а также сагиттального гребня черепа. Вряд ли можно сомневаться в том, что ведущим в этом процессе эволюции хищных млекопитающих было изменение инстинктов животного, переходившего от питания насекомыми к добыче и использованию более крупных животных. Первым результатом такого перехода могла быть только усиленная тренировка челюстных мыши с ее последствиями — увеличением их массы (а также их кровоснабжения) и увеличением поверхности их прикрепления на частях скелета. Мы знаем, что такие изменения могут иметь характер приспособительных модификаций (опыты Хюртля) и не лишены целостности. Однако в процессе эволюции несомненно происходила стабилизация таких изменений (через элиминацию особей с недостаточным развитием челюстей и их мускулатуры, случайно выросших в благоприятных условиях питания мелкими животными) на почве индивидуальной конкуренции в добывании крупных животных. Соответственно происходила «фиксация» новой нормы (при этом менялся и способ ее развития — в автономном процессе формирования мышцы ее объем определяется числом волокон, а не их толщиной, как при тренировке). Однако в то же самое время известные преимущества в борьбе за существование имели и те особи, которые обладали наследственно еще более совершенной нормой и, в частности, особи с наследственными изменениями в зубной системе (которая, как известно, от «упражнения» заметно не меняется, во всяком случае форма и величина зубов не модифицируются). Поэтому и ведущая форма отбора имела здесь бесспорное значение.

Рис. 12. Посевы Camelina linicola и С. glabrata, сделанные одновременно и выращенные в совершенно одинаковых условиях. Видно наличие наследственных различий нормы реакций. Вверху — С. linicola, внизу — С. glabrata. По Н. В. Цингеру, 1909
Значение приспособительной модификации, как переходного этапа в процессе эволюции, играющего в течение первоначального периода формирования новой адаптивной нормы (или замены главной нормы добавочной) руководящую роль, ограничивается скоростью этих изменений. Эта скорость так значительна, что при быстрой смене условий внешней среды приобретает характер решительного скачка, сразу переносящего организм в новую среду. Мы уже отмечали скорость этого процессса на примерах горной формы Capsella bursa pastoris. To же относится к модификации ярового рыжика Camelina glabrata, приведшей в посевах льна, т. е. за время развития этой культуры, к образованию льняного рыжика С. linicola. Образование высокого стебля с длинными междоузлиями и узкими листьями, возникшее первоначально как модификация (как это наблюдается при высеве С. glabrata в посевах льна), здесь уже стабилизировалось, т. е. наследственно «зафиксировано». С. linicola дает высокие стебли с узкими листьями и вне посевов льна (рис. 12, 13). С другой стороны, увеличение размера семян представляет результат прямого отбора в его ведущей форме (Н. В. Цингер, 1909).
Конечно, этот быстрый переход возможен лишь на основе, подготовленной предшествующей эволюцией организма. Ведь сама способность организма к адаптивной модификации является частью унаследованной нормы его реакций, выработанной ведущей формой отбора.
Это особенно ясно в случаях резко выраженного полиморфизма, т. е. существования нескольких целостных адаптивных норм. У большинства высших растений поверхность листьев, их строение, опушение, толщина кутикулы и численность устьиц легко подвергаются явным модификациям в зависимости от интенсивности освещения, так что говорят о световых и теневых листьях. В частности, обе реакции хорошо выражены у вербейника (Lys-machia vulgaris). Однако теневые формы Lysimachia numullaria и Dactylis glomerata сохраняли свои характерные признаки и при длительной культуре (многие годы) на солнечном месте (Turesson, 1922). Здесь световая модификация оказалась утерянной, а теневая стабилизировалась.
Иногда и длительная история организма в новой среде не исключает возможности частичного возврата к исходным формам при восстановлении прежних условий. Так, по исследованиям Н. Ва-куленко (1940), при прорастании зимних почек типичного водяного растения Myriophyllum verticillatum в воздушной среде (во влажной камере) на верхних частях стебля развивались еще цельнокрайние листья. Здесь совершенно ясен переход от амфи-биотического растения с двумя нормами (причем первично главной нормой была наземная, а добавочной — водяная) к чисто водяному растению, почти уже утратившему свою прежнюю наземную норму и стабилизировавшему развитие своей новой нормы.

Рис. 13. Сравнение Camelina linicola с различными модификациями С. glabrata. Нормально льняной рыжик растет в густых посевах льна. 5 — С. linicola, выращенный в редком посеве; 6 — С. glabrata, выросший в тех же условиях, что и С. linicola на рис. 5. Различия между 5 и б имеют генотипический характер; 7 — С. glabrata, развивавшийся в посеве льна, соответственно обычным условиям развития С. linicola; 8— С. glabrata, развившийся на просторе. Оба последних экземпляра получены из семян одной особи. Различия между 7 и 8 покоятся на модификациях. Внизу показаны 4-й, 6-й, 9-й, 12-й и 15-й листья соответствующих растений. По Н. В. Цингеру, 1909
Такой переход реализуется, конечно, с большой легкостью и скоростью при подходящих для этого внешних условиях, когда те факторы, которые раньше имели лишь локальное или временное значение, становятся постоянными для данного вида организмов. Изложенное представлено на рис. 14, А, В, С, Д.

Рис. 14. Отбор в разнообразных и колеблющихся условиях внешней среды (с учетом различных форм индивидуального реагирования, т. е. модификаций). А — отбор при установившихся (в данном многообразии) условиях (стабилизирующая форма отбора). Вариационные кривые: 1 — всех особей; 2 — особей, вышивающих в реально встречающихся различных условиях (условно «нормальные» особи, условно благоприятные и частично благоприятные уклонения); 3 — особей, выживающих и оставляющих потомство в обычной средней «нормальной» обстановке. Точками обозначена зона безусловной элиминации уклонений, вредных при всех условиях (мутаций и морфозов). Косой штриховкой обозначена зона вариантов, элиминируемых при обычных, нормальных условиях, но выживающих в известных уклоняющихся, но реально встречающихся условиях. В — отбор при изменении условий в определенном направлении (с сохранением их многообразия). Ведущая форма отбора. Вариационные кривые: 1 — всех особей; 2— особей, выживающих в различных условиях новой среды; 3 —особей, выживающих в новой «нормальной» обстановке. С, D — другой 'план изображения механизма стабилизирующей и ведущей форм отбора.
Эволюция половых признаков — одна из интереснейших биологических проблем, до настоящего времени еще далеко не разрешенная. Своеобразие этой эволюции обратило на себя внимание Ч. Дарвина, отметившего многие противоречия тем закономерностям, которые были им установлены для общей организации живых существ. Дарвину пришлось ввести особый принцип полового отбора для объяснения части этих противоречий. Однако тот же Дарвин обратил внимание и на то обстоятельство, что многочисленные факты передачи вторичных половых признаков с самца на самку противоречат и этому принципу. Относящиеся сюда факты не получили своего объяснения и до настоящего времени. Я здесь не принимаю во внимание физиологических объяснений передачи самцовых признаков предположениями о повышении концентрации мужского гормона у самки. Если бы они и оказались в известных случаях верными, то, вскрывая механизм этой передачи, они все равно потребовали бы раскрытия причин исторического возникновения этого механизма. Дело касается, однако, не только вторичных половых признаков. И в эволюции копулятивных органов, их вспомогательных частей и даже самих половых желез мы можем отметить немало явлений, непонятных с обычных точек зрения.
Одним из важнейших доводов в пользу эволюционного учения были в свое время факты наличия у эмбрионов или личинок зачаточных органов провизорного значения, которые не достигают полного развития у данного организма, но вполне сравнимы с зачатками дефинитивных органов других организмов. Такие зачаточные органы мы считаем унаследованными от предков, у которых они были вполне развиты. Это — явления так называемой рекапитуляции предковых состояний.
Однако в половой системе мы находим зачатки подобного же характера, которые не могут быть обозначены как рекапитуляция. У самца млекопитающих в течение эмбрионального периода развиваются яйцеводы и зачаточная матка (uterus masculinus), а у самки развивается копулятивный орган, превращающийся позднее в клитор. Считается, что половая система развивается на некоторой «индифферентной» основе, общей для обоих полов. Эта основа, однако, вовсе не индифферентна: она попросту совмещает дифференцированные признаки обоих полов. Млекопитающие не произошли от гермафродитов — их предки были раздельнополыми. Следовательно, в этом совмещении зачатков органов обоего пола нет рекапитуляции. Самцы никогда не обладали яйцеводами и маткой, а самки не обладали копулятивным органом. Их зачатки не являются у них повторением предковых состояний. Наоборот, они возникают сначала у одного пола, затем как эмбриональные признаки у другого пола, и, наконец, переносятся в большей или меньшей мере и на взрослое животное. Можно было бы думать, что это связано с самим механизмом наследования, в котором участвуют оба пола. Однако и это не так просто. Механизм раздельного наследования половых признаков имеется с самого начала их появления — половые признаки возникают именно у одного из полов. Только в дальнейшей эволюции раздельное наследование нередко почему-то нарушается.
Это нарушение, ведущее к передаче признака на другой пол, иногда определяется выгодой и связано с действием естественного отбора (например, передача органа защиты — рогов — с самца на самку или передача покровительственной окраски самки на самца). Такие случаи разобраны Дарвином. Однако они не составляют общего правила. Наоборот, гораздо чаще наблюдается другое: передача признаков с одного пола на другой не связана с ясной выгодой и поэтому вряд ли объяснима действием обычного естественного отбора. Во многих случаях, конечно, трудно судить о биологическом значении даже таких признаков, как рога самки. Не всегда они имеют значение органов защиты. Однако нередко мы не только не видим положительного значения передачи полового признака с одного пола на другой, но можем даже предполагать, что эта передача скорее неблагоприятна. Для вторично-половых признаков это иногда кажется почти бесспорным.
В самом деле, если вторично-половой признак, например самца, имеет известное биологическое значение (привлечение и возбуждение самки, стимуляция нормального полового цикла у самки и, в результате этого, обеспечение спаривания и оплодотворения), то с передачей его на самку он тем самым теряет свое значение у самца, т. е. обесценивается. С другой стороны, самка приобретает ненужные ей органы (например мозоли на задних конечностях у некоторых амфибий, шпоры на ногах у некоторых куриных), теряет свои средства пассивной защиты (покровительственную окраску) и приобретает яркую, бросающуюся в глаза окраску, делающую ее более легкой добычей хищников. Приобретение самкой яркого оперения, как правило, не только биологически бесцельно, но и прямо вредно. Поэтому в процессе эволюции «независимых» половых признаков самца у самки вырабатывается особый тормозящий механизм, противодействующий этой передаче. И все же передача нередко осуществляется вопреки наличию такого механизма.
Нередко такая передача признака с самца на самку имеет резкий, скачкообразный характер — она не подготовляется в процессе медленной эволюции самки. И в этом мы видим своеобразие, нуждающееся в объяснении. И, наконец, соответственно двум разным механизмам индивидуального развития вторично-половых признаков самца — как зависимых от мужского полового гормона или от него независимых (с торможением женским гормоном у самки) — имеются и два разных способа передачи сам-цовых признаков с самца на самку: через повышение чувствительности тканей к мужскому половому гормону или через понижение их чувствительности к женскому гормону (Б. Г. Новиков, 1940). Два разных пути ведут к одному и тому же результату. Очевидно, в основе этих явлений лежат какие-то более сложные биологические закономерности, а не простые физиологические процессы, связанные с механизмом определения пола.
Однако обычное объяснение исторических закономерностей направлением естественного отбора, идущим на основе селекционного преимущества известных уклонений (в данном случае самок, обладающих самцовыми признаками), очевидно, также недостаточно.
1. Передача распознавательных вторично-половых признаков с одного пола на другой. Основные факты, касающиеся явления передачи самцовых признаков самкам, собраны уже у Ч. Дарвина. У оленей, как правило, рога имеют только самцы. Только у северного оленя самка обладает хорошо развитыми рогами. Однако Дарвин отмечает, что у самок некоторых видов оленевых постоянно или случайно встречаются следы рогов. Например, большинство самок вапити (Cervus canadensis) на месте рогов имеет острый костяной выступ. У антилоп имеется полный ряд форм — от видов с рогами только у самцов, через такие виды, у которых самки имеют ничтожные зачатки рогов или очень небольшие рога, до видов с самцами и самками, несущими одинаковые рога. У некоторых животных рога имеют обычно и самцы, и самки, однако у самцов они, как правило, значительно больших размеров (козлы, бараны, быки). При кастрации самца его рога получают особенности рогов самки. Ч. Дарвин отмечает вообще различное влияние кастрации самца на развитие рогов у оленей и у полорогих жвачных.
У фазанов самцы очень красивы и резко отличаются от самок. Однако у ушастых фазанов «прекрасные хвостовые перья, большие хохлы на ушах и ярко-красный бархат вокруг головы свойственны обоим полам» (Дарвин). У обыкновенного павлина (Pavo cristatus) шпоры имеются только у самца. Однако у яванского павлина (P. muticus) шпорами обладает и самка. Из рептилий резкие половые различия бывают у хамелеонов. У мадага-скарского Chamaeleon bifurcatus на голове самца имеется пара больших, направленных вперед рогообразных выростов. Однако у этого вида и самка обладает небольшими зачатками таких выростов.
Бабочки-голубянки (Lycaena) могут служить примером иногда ясно выраженных половых различий. «У Lycaena agestis оба пола имеют крылья бурые, окаймленные оранжевыми глазчатыми пятнами и сходные между собой. У L. oegon крылья у самцов прекрасного голубого цвета с черной каймой, а у самки — бурые, с такой же каймой и очень похожие на крылья L. agestis. Наконец, у L. arion оба пола голубые и схожи, хотя у самок края крыльев несколько темнее и черные пятна проще, а у одного индийского ярко-голубого вида оба пола еще более похожи друг на друга» (Дарвин). Здесь, следовательно, голубая окраска была сначала приобретена самцом, а затем передана и на самку. У кузнечиков звуковые органы функционируют только у самцов, однако в зачаточном виде они встречаются иногда и у самок. У цикад, как правило, и самки обладают звуковым аппаратом, но он у них не функционирует. У жуков из Lamellicornia самцы обладают различными рогообразными выростами. Однако иногда зачаточные выросты наблюдаются и у самок. Иногда встречается такая же бесполезная передача признака в обратном направлении — с самки на самца. Дарвин приводит следующий замечательный пример такой передачи: «У пчел аппарат для собирания пыльцы употребляется в дело одними самками, собирающими пыльцу для личинок; и, тем не менее, у большинства видов он отчасти развит и у самцов, которым совершенно бесполезен, и развит очень хорошо у Bombus или шмеля».
В результате своего обзора половых признаков Ч. Дарвин приходит к некоторым выводам, имеющим фундаментальное значение для всей этой проблемы. Во-первых, отмечается позднее развитие и значительная изменчивость половых признаков по сравнению с видовыми признаками. Во-вторых, указывается, что перенос самцового признака на самку и, следовательно, превращение полового признака в видовой сопровождается развитием признака в более раннем возрасте. Дарвин пишет относительно рогов северного оленя: «На основании этих различных фактов мы можем заключить, что обладание хорошо развитыми рогами у самки северного оленя зависит от того, что самцы приобрели их первоначально для поединков с другими самцами, и стали они развиваться у них, вследствие какой-то неизвестной причины, в необычайно раннем возрасте, вследствие чего они передались обоим полам».
2. Скачкообразный характер передачи. В настоящее время накопилось довольно много фактов, показывающих внезапный, скачкообразный характер передачи признака с одного пола на другой. Среди упомянутых уже голубянок известны виды (L. argiodes, L. orion), у которых наравне с исходными бурыми самками встречаются и самки, обладающие самцовой голубой окраской. Среди бабочек североамериканского рода Epicalia известны 9 видов с исключительно яркими самцами, но одноцветными самками, и 2 вида с самками почти столь же яркими, как и самцы. У южноамериканской бабочки Colias lesbia, кроме обычных бледно окрашенных самок, встречаются самки с более темными оранжевыми крыльями самцового типа. У жука-плавунца (Dytiscus marginalis) самки нормально обладают надкрыльями с продольными бороздками (как у родственных жуков). Однако встречаются иногда и самки с гладкими надкрыльями самцового типа. У некоторых близких видов из средней Европы такие самки столь же обычны, как и «нормальные». Иногда развитие и передача признака стимулируются климатическими влияниями. Так, у упомянутых плавунцов (Dytiscus) самцовые гладкие надкрылья переходят на самку главным образом в теплых районах Европы. У камбалы (Pleuronectes platessa) самцы обладают в Ла-Манше (Английский канал) немногими чешуями с шипами; северне'е эти признаки выражены резче, и еще резче они выступают в Балтийском море. В североамериканских районах Арктики ее место занимает близкий вид PI. glacialis, у которого чешуи с колючками являются ярким самцовым признаком. Однако у берегов Аляски и самки того же вида обладают колючками (Cunningham, цит. по Winterbottom, 1929).
Из рептилий голубое горло самцов ящериц Lacerta viridis и L. laevis встречается иногда и у самок. Зеленые бока самцов ящериц L. agilis встречаются иногда и у самок. Носовые рогообразные выросты, характерные для самцов многих видов хамелеонов, имеются и у самок Ch. bitaeniatus (Guenoi;, 1925, цит. по Winterbottom, 1929) и Ch. matschiei (Meisenheimer, 1921).
У птицы фрегата (Fregata aquila) оба пола имеют черное оперение, подобное самцам других видов — F. magnificans и F. minior, У палестинского воробья (Petronia brachydactyla) птенцы и взрослые птицы обоего пола имеют скромную серовато-бурую окраску. Птенцы и самки домашнего воробья (Passer domesticus) сохранили это примитивное оперение, но самец окрашен более ярко. У полевого воробья (P. montanus) оба пола приобрели эту более яркую самцовую окраску. У огромного большинства видов колибри самцы отличаются от самок значительно более яркой окраской. Однако у видов Agirtia, Cclibri, Panterpe блестящая окраска самцов перешла и на самок. У индийских фазанов Bambasicola и Galloper-dix в пределах одних и тех же видов встречаются самки без шпор и самки со шпорами. Между тем, у кур Зондских островов Rhizo-thera и Acomus самки постоянно обладают такими же шпорами, как и самцы. У кускуса (Phalanger maculatus) из сумчатых Новой Гвинеи самки имеют серую однотонную окраску, а самцы рыжие с белыми пятнами. Однако на острове Вайгео, к северу от Новой Гвинеи, самки того же вида имеют самцовую окраску с белыми пятнами. У американской вилорогой антилопы (Antilocarpa) 20% самок обладают рогами. У азиатского муфлона самки безроги.
У европейского муфлона на Сардинии самки обычно безроги, а на Корсике имеют рога. У других овец рога имеются у самок наравне с самцами. У современных жираф рогообразные выросты имеются у обоих полов. Однако у ископаемых Samotherium boissieri и Palaeoiragus rouenii рога были только у самцов. У всех почти оленей рога имеются только у самцов. У современного северного оленя (Rangiber tarandus) и самки обладают хорошо развитыми рогами. Однако это совсем недавнее приобретение самок. В Саянах встречается еще исходная форма северного оленя с безрогими самками (А. Машковцев, 1940). Л в начале 19-го века и в Европейской России встречались еще безрогие самки северного оленя в Казанской (?) губернии, по указанию Meisenheimer; вероятно, это относилось к красноуфимскому стаду Пермской области.
3. Механизм передачи признаков с одного пола на другой. Современные экспериментальные исследования показали, что механизм развития половых признаков может быть различным и соответственно различным оказывается и механизм передачи признаков с одного пола на другой. Развитие половых признаков может быть обусловлено более или менее непосредственно генетическими и морфогенетическими факторами в узком смысле (половые различия беспозвоночных и в значительной мере первичные половые признаки позвоночных), а может быть обусловлено более или менее сложным сочетанием этих факторов с гормональными влияниями (первичные и в особенности вторичные половые признаки позвоночных). В развитии самцовых признаков позвоночных обнаруживается их явная зависимость от мужского полового гормона («зависимые» самцовые признаки, по М. М. За-вадовскому) или же они развиваются независимо от этого гормона («независимые» признаки, по М. М. Завадовскому, 1922). В последнем случае их развитие подавляется у самки вследствие противодействия со стороны женского гормона.
Передаются, как мы видели, с одного пола на другой различные половые признаки, как зависимые в своем развитии от полового гормона, так и независимые, как связанные в своем происхождении с половым отбором, так и не связанные ни с подготовкой к спариванию, ни с брачным сезоном. Передача возможна не только с самца на самку, но и в обратном направлении (покровительственная окраска самки, подражательная окраска у бабочек, аппарат для собирания пыльцы у пчел и шмелей, сосцы у самцов млекопитающих).
Механизм этой передачи исследован, по-видимому, только у птиц. В новейшее время вопрос этот разрабатывался Новиковым (1940), с одной стороны, на воробьиных птицах и, с другой, на некоторых куриных. У обыкновенного щегла (Carduelis carduelis) и самец и самка обладают исходной светлой «роговой» окраской клюва. У домашнего воробья (Passer domesticus) и у зяблика (Fringilla coelebs) самцы приобретают в брачный период темную окраску клюва. Эта окраска зависит в своем развитии от мужского полового гормона и может быть индуцирована также у самки путем его инъекции. Однако у вьюрка (F. montijringilla) брачная темная окраска клюва развивается весной и летом у обоих полов. Признак перешел на самку, но сохранил сезонный характер. Он сохранил и свою зависимость от мужского полового гормона. Перенос признака на самку связан с повышением чувствительности тканей к мужскому половому гормону (который у самки продуцируется клетками медуллярной зоны яичника). Эксперименты показали, что для индукции брачной окраски клюва у вьюрка требуется в 4 раза меньшая доза гормона, чем у зяблика. У полевого воробья (P. montanus) черная окраска клюва также имеется у обоих полов, но сезонность этого признака почти утрачена — в период максимальной депрессии гонад черный пигмент исчезает только у основания клюва. Переход к постоянному мономорфизму происходит у полевого воробья за счет дальнейшего повышения чувствительности тканей к половому гормону — для индукции черной окраски клюва у кастрата требуется в 10 раз меньшая доза тестостерона, чем у домашнего воробья. Наконец, у близких к воробьям снегирей (Pyrrhula purrhula) черная окраска развита у обоих полов и не подвергается сезонным изменениям. Половой признак окончательно превратился в постоянный видовой признак.
Вместе с повышением чувствительности тканей к мужскому половому гормону реакция происходит с большой легкостью и фактически теряет свою типичную зивисимость от наличия гормона. У полевого воробья реакция почернения клюва оказывается трудно обратимой — через 1 — 1,5 года после кастрации кончик клюва сохраняет еще следы темной окраски. У снегирей же черная окраска клюва развивается даже после кастрации молодых птиц, т. е. как-будто независимо от мужского полового гормона (вероятно, индуцируется заранее, а после детерминации осуществляется, как необратимый процесс).
Во всяком случае, передача признака с самца на самку связана в разобранном ряду форм с повышением реактивности тканей на мужской половой гормон. Очевидно таковым же является и механизм передачи на самку других самцовых зависимых признаков, как головной убор и шпоры куриных, а также рога млекопитающих. И здесь передача связана с повышением чувствительности тканей, т. е. со снижением порога реактивности тканей на мужской половой гормон. Когда этот порог достигает уровня нормальной концентрации мужского гормона у самки, половой признак сразу (скачкообразно) переносится на самку.
Развитие и передача самцового оперения на самку обусловлены у куриных, а вероятно и у многих других птиц, несколько иным механизмом. В этом случае развитие самцовых признаков — «независимое» от мужского полового гормона, но подавляется у самки действием женского полового гормона. В случае переноса самцового оперения на самку это оказывается результатом снижения реактивности тканей на овариальный гормон. Новиков показал, что у серой куропатки (Perdix perdix), которая, по Дарвину, обладает вторичным мономорфизмом, т. е. самцовым оперением у обоих полов, можно путем инъекции больших доз овариального гормона добиться развития более примитивного оперения ювенального типа.
У большинства диморфных форм с ювенальным оперением сходно именно оперение самки, а не самца. Самец обладает, как правило, более высоко дифференцированными оперением и его окраской. Поэтому можно вывести заключение, что у серой куропатки передача самцового оперения на самку вызвана снижением чувствительности тканей (т. е. повышением порога их реактивности) к овариальному гормону. При экспериментальном повышении концентрации последнего развитие самцового оперения еще может быть подавлено, и тогда проявляется вновь исходное примитивное оперение самки( сходное с ювенальным). Таким образом, Новикову удалось доказать, что серая куропатка действительно произошла от диморфной формы вследствие переноса самцового оперения на самку (и превращения полового признака в видовой). То же самое относится к частичному мономорфизму самцового типа у тетерева (Lyrurus tetrix), именно к реализации металлической окраски перьев спины, а также к развитию самцового зеркальца на второстепенных маховых перьях у самок руанской утки (дикой окраски). Из диких уток таким зеркальцем обладает только самка кряквы. У других видов уток — это характерный самцовый признак. Развитие зеркальца у руанских уток может быть заторможено введением дополнительной дозы овариального гормона (цит. по Б. Новикову, 1940).
С другой стороны, в случаях также несомненного вторичного мономорфизма у каменных куропаток (Новиков), голубей и у чибисов подобные же опыты не привели ни к какому результату. Очевидно, у этих птиц ткани уже полностью утратили способность к специфической реакции на женский половой гормон. Самцовое оперение окончательно превратилось в независимый видовой признак.
Таким образом, перенос самцовых «независимых» признаков на самку связан с постепенным снижением реактивности (т. е. повышением порогового уровня) тканей на женской половой гормон, который нормально препятствует развитию самцовых признаков (Новиков).
Соответственно наличию двух механизмов развития вторичных половых признаков у птиц и передача самцовых признаков на самку осуществляется двумя различными путями: 1) «зависимые» признаки самца передаются через увеличение чувствительности к мужскому гормону, т. е. через снижение нижнего порога реактивности тканей до уровня нормальной концентрации мужского гормона у самки; 2) «независимые» самцовые признаки передаются через уменьшение чувствительности к овариальному гормону, т. е. через повышение нижнего порога реактивности тканей до уровня нормальной концентрации этого гормона у самки.
Оба как-будто противоположных процесса заключают в себе одинаковое биологическое содержание — они оба ведут к' большей стабильности самцовых признаков у самца, они делают их проявление независимым от случайных колебаний в концентрации полового гормона, а через это и от случайных колебаний во внешних факторах, определяющих периодичность его продукции (температура, свет).
Оба процесса имеют биологический смысл только у самца. Однако они могут привести в известной фазе своего развития к изменению признаков самки. Интересно, что эта передача самцовых признаков на самку вовсе не обязательно связана с гормональным механизмом развития признаков пола. Мы видели, что такая передача наблюдается и у насекомых. Это объясняется, очевидно, тем, что весь механизм определения пола и развития его признаков всегда почти основан на чисто количественных различиях в одинаковых факторах (в концентрации одних и тех же половых гормонов, тех же морфогенных веществ и в простой или двойной дозе наследственных факторов половой хромосомы), а не на качественных различиях. Поэтому при известной степени изменения порога нормальной для одного пола реактивности тканей (ведущего к большей стабильности признаков у одного пола) достигается осуществление этой реакции; и у другого пола. Так, на известном уровне повышения чувствительности к мужскому гормону или снижения чувствительности к женскому гормону (ведущих к устойчивости в проявлении половых признаков у самца) происходит реализация полной самцовой морфогенетической реакции у самки.
Стабилизация признаков одного пола, конечно, не всегда доходит до того уровня, когда эти признаки переносятся на другой пол. В случае явной вредности этих признаков для самки, этой передаче будет препятствовать систематическая элиминация самок, обладающих самцовыми признаками. Установится некоторый предельный «оптимум» стабильности признаков самца. Дальнейшая стабилизация прекратится. Наконец, в некоторых случаях половой признак может вторично развиться из видового, и тогда весь механизм его развития оказывается иным и может имитировать передачу признака на другой пол. Возможно, что таковым был путь развития рогов у антилоп. Дж. Гексли считает, что они первоначально возникли у обоих полов, а позднее становятся самцо-вым признаком через подавление их развития у самок. Во всяком случае, бросается в глаза, что у антилоп, в отличие от оленей, рога у большинства видов имеются у обоих полов и лишь у немногих видов только у самцов. Кроме того, в отличие от типичной картины скачкообразного «переноса» признака с одного пола на другой, здесь наблюдаются все переходы от рогатых самок к безрогим.
4. Передача других половых признаков с одного пола на другой. Перенос признаков одного пола на другой не ограничивается одними только распознавательными вторично-половыми признаками. Этот процесс распространяется и на органы заботы о потомстве, на вспомогательные органы спаривания и даже на первичные половые признаки — на копулятивные органы и на половые железы. Много фактов такого рода собрано в известной книге Meisenheimer (1921). И у беспозвоночных наблюдается передача особенностей половых частей с одного пола на другой. Так, например, у раков встречаются добавочные половые отверстия, соответствующие половым отверстиям другого пола (самка имеет нормальное отверстие у основания третьей пары абдоминальных ножек, а самец — у основания пятой пары). У самца речного рака передняя пара абдоминальных ножек служит аппаратом для переноса спермы. У самки эти ножки, как правило, рудиментарны. Однако иногда они развиваются довольно значительно и тогда приобретают типичную полутрубчатую структуру ножек самца и отличаются от них лишь несколько меньшей величиной. Мы уже отмечали факты развития сосцов и даже зачаточных молочных желез у самцов млекопитающих (включая человека). Число этих органов обычно соответствует их числу у самок. Это, конечно, не объясняется унаследованием примитивного состояния, даже если допустить, что у исходных предков млекопитающих оба пола участвовали в выкармливании детенышей. Переносятся на самца не какие-либо примитргвные формы железистого поля (как у однопроходных), а именно зачатки высоко развитых желез и сосцов плацентных млекопитающих. На этой базе уже вторично млечные железы могут достигнуть полного развития у самца и могут быть использованы самцом для выкармливания детенышей. Эта высшая фаза передачи млечных желез на самца достигнута некоторыми летучими мышами (Cynorycteris grandidieri с Занзибара и Супоpterus marginatus с Цейлона). В этом случае самка рождает двух детенышей, из которых один выкармливается самкой, а другой — самцом. В данном случае передача признака с самки на самца начинается с эмбрионального зачатка и завершается затем полным развитием признака у взрослого животного. С тем же порядком передачи мы встречаемся и при рассмотрении копулятивных органов и первичных половых признаков. Он явно доказывает вторичность этого процесса.
В этом случае мы сталкиваемся с целым рядом совершенно непонятных явлений. У самцов Diemyctylus viridenscens имеются на нижней стороне бедер мозолистые органы, служащие для захватывания самки при копуляции. Однако подобные же органы развиваются в ослабленной степени и у самок. Бедренные органы ящериц, имеющие сходное назначение, встречаются в ослабленном выражении и у самок некоторых Lacertidae и Iguanidae (Meisenheimer). Даже настоящие парные копулятивные органы ящериц имеются в зачаточном виде и у самок. Непарный penis имеется и у самок крокодилов и черепах. Он небольшой, но обладает таким же строением, как у самцов. У Trionix он так же, как у самца, имеет на конце 5 выступов. И у птиц, обладающих копулятивным органом, таковой всегда имеется и у самки, но только меньших размеров. Наиболее замечательные явления относятся, однако, к млекопитающим.
У самок млекопитающих у переднего конца половой щели постоянно развивается зачаточный копулятивный орган, называемый клитором. Его строение сходно с копулятивным органом самца — пенисом. Имеется пара основных эректильных тел — corpora cavernosa clitoridis, которые прилегают к внутренней стороне тазовых костей и, сходясь впереди, образуют непарное пещеристое тело в стволе клитора. На переднем конце последнего имеется непарное тело головки — corpus cavernosum glandis. Имеется и пара уретральных пещеристых тел, но у самки они позади широко расходятся, охватывая мочеиспускательный канал (так как последний у самки не всегда имеет отношение к клитору). Все эти тела развиты у самки слабее, чем у самца, но все же способны к эрекции и, следовательно, не лишены функционального значения. Строение копулятивного органа разнообразно у разных млекопитающих и также разнообразно строение клитора. У эхидны конец пениса разделяется на четыре стволика с отдельными головками. То же разделение имеется и на клиторе самки. У сумчатых часто имеется раздвоенный пенис, и в этих случаях и клитор оказывается раздвоенным (особенно у Didelphys и Perameles — двойной пенис и двойной клитор). Однако у кенгуру (Macropodidae) имеется простой пенис и соответственно простой клитор.
В тех случаях, когда у млекопитающих (например у хищных) имеется кость в копулятивном органе — os priapi — соответственно имеется кость такой же формы (но меньшей величины) и в клиторе.
Сходство клитора с копулятивным органом увеличивается с переносом мочеиспускательного канала — уретры —- на клитор. Все фазы этого процесса могут быть прослежены у грызунов. У белки и у бобра уретра открывается еще далеко от клитора. У дикобраза (Hystrix) отверстие уретры лежит у основания клитора и продолжается в бороздку, образованную расщепленным препуциумом. У многих мышей препуциальная бороздка замыкается и образует канал, переносящий отверстие мочеиспускательного канала на самый конец (головку) клитора (Mas decumanus и другие мыши). Сходные отношения с мочеиспускательным каналом в клиторе имеются у крота, землеройки, у летучих мышей и почти у всех приматов. Половая щель и влагалище остаются, однако, позади клитора. Иногда при этом наблюдается огромная величина клитора. Таков, например, клитор полуобезьян Stenops, Lemur varius и Octoclinus crassicaudatas. Еще выше развит клитор у некоторых обезьян Нового Света — Mycetes и, в особенности, Ateles. Клитор здесь вполне сходен с копулятивным органом самца, но обладает снизу бороздкой, переходящей позади в половую щель. Однако в данном случае клитор мало эректилен. Большой клитор бывает и у хищников (например Cryptopracta): он содержит os priapi, выступает иногда далеко вперед, снабжен продольной бороздкой, а снаружи покрыт, как и пенис, шипами.
Наконец, совершенно удивительные отношения устанавливаются у гиен. Здесь сходство клитора с копулятивным органом достигает своего высшего предела. У пятнистой гиены Hyaena crocu-ta (по Watson, 1877, и Matthew, 1939, см. у Meisenheimer, 1921, и у Huxley, 1938) оба пола внешне вообще неотличимы. Клитор лежит в кожной складке и способен к полной эрекции подобно пенису. Он обладает хорошо выраженным препуциумом и головкой. Он обладает также объемистыми пещеристыми телами и мышцей — ретрактором, втягивающим его назад внутрь кожной сумки. Половой щели (vulva) в обычном смысле не существует. Клитор пронизан вдоль идущим мочеполовым синусом (преддверием влагалища) и, следовательно, половая щель представлена отверстием в конце клитора. Копуляция происходит через этот канал в клиторе и поэтому вообще чрезвычайно затруднена (Matthew, 1939). Через него же (т. е. через клитор) происходит и деторождение. Все это устройство поражает своей нецелесообразностью.
Иногда у самок млекопитающих развивается и мошонка. Эмбрионально ее закладка имеется всегда, но обычно она не развивается далее и преобразовывается лишь у некоторых млекопитающих (и у человека) в большие губы. У самок широконосой обезьяны Mycetes большие губы имеют вид вздутых полушаровидных валиков, наполненных жировой тканью. Неровно-складчатая поверхность кожи увеличивает сходство с мошонкой. У самок хищника Cryptoprocta кроме большого клитора имеется также настоящая по внешности мошонка в виде пары округлых вздутий, покрытых длинными волосами. Наконец, у описанных уже самок пятнистой гиены (Hyaena crocuta) на том же месте промежности, как у самца, имеется неотличимая от настоящей мошонка в виде парного вздутия, покрытого нежными бурыми волосами, совершенно как у самца. Конечно, внутри мошонки вместо семенников имеется лишь жировая ткань. Как уже было сказано, в данном случае по виду наружных половых органов вообще нельзя отличить самца от самки.
Однако у млекопитающих перенос половых признаков с одного иола на другой не ограничивается наружными половыми органами. Этот процесс распространяется и на внутренние части полового аппарата — на половые пути. И у самки развиваются части мужских выводных путей, в том числе зачаток придатка семенника, образующий epoophoron, а также и некоторые придаточные железы, образующие у самки железы преддверия влагалища. Однако наиболее замечательна передача признаков в обратном направлении — с самки на самца.
Мюллеровы каналы, как зачатки женских половых путей (яйцеводов, матки и влагалища), являются специфическими органами самки, однако они закладываются и у самца. Нередко их остатки сохраняются и у взрослых самцов млекопитающих. У насекомоядных, у многих хищных и у приматов на внутреннем конце мочеполового протока имеется слепой выступ, впадающий в этот проток — vesicula prostatica. У бобра (Castor fiber) этот пузырек разделен на две ветви и вполне соответствует двурогой матке самки. Эта «мужская матка» (uterus masculinus) обнаруживает чрезвычайные различия у близких видов и даже значительные индивидуальные различия в пределах известного вида. Так, у канадского бобра (С. canadensis) не заметно ни следа «мужской матки». У половины особей домашнего козла «мужской матки» нет вовсе, у других — небольшой пузырек; иногда же имеется типичная двурогая матка.
У дельфина Phocaena communis наблюдаются у самца все переходы от небольшого полового выступа мочеполового протока до типичной двурогой матки. Велики индивидуальные различия и у человека. Часто это — удлиненный пузырек, обычная vesicula proslatica без добавочных частей, иногда же — настоящая матка (с маточными железами) до 12 см длиной, с яйцеводами и даже ostia и fimbria по их краям, с шейкой матки и влагалищем. Наравне с этим имеется совершенно нормальный мужской аппарат, так что в этих случаях пет речи об интерсексуальности («гермафродитизме»).
Передача таких образований на другой пол необъяснима с обычных точек зрения — в ней нет биологического смысла. Не только нет выгоды для вида в целом, но иногда можно предполагать даже наличие известной вредности (как в случае, описанном для пятнистой гиены). Обычная, ведущая форма естественного отбора не могла привести к таким результатам. «Объяснение» Meisenheimer, ссылающегося на «силу наследственности» и на «силу передачи», исходящие от другого пола, также не выдерживает критики.
Для того чтобы представить себе причины переноса половых признаков на другой пол, рассмотрим сначала основные особенности, отличающие механизм индивидуального развития половых признаков от факторов индивидуального развития других организационных (в том числе «видовых») признаков, присущих обоим полам.
5. Факторы, определяющие перенос половых признаков с одного пола на другой. Факторы индивидуального развития половых признаков сходны с обычными факторами развития всей организации, и мы можем вообще говорить о генетических, морфогенетических и гормональных факторах. Первые общи всем клеткам и не имеют локализирующего значения. Они обусловливают явление специфического (для данного вида организмов) самодифференцирования материала (вместе с его видовыми, расовыми и даже индивидуальными особенностями) через локализирующие влияния внешних факторов, различий в плазме яйца, имеющихся уже в яйце и развивающихся позднее, количественных различий типа градиентов, а также благодаря различиям количественного и качественного характера, устанавливающимся через взаимодействия частей в морфогенетических системах. Вторые, т. е. морфогенетические факторы, имеют в основном локализирующее значение и создаются в процессе развития, начиная с образования яйца, его оплодотворения, дробления, гаструляции и в течение развития зародыша, при непрерывном взаимодействии с внешней средой, на базе небольших вначале различий, путем перемещения и взаимодействия развивающегося материала. Третьи, т. е. гормональные факторы, могут иметь лишь стимулирующее или тормозящее значение для дифференцированных уже процессов, развертывающихся на основе взаимодействия генетических и морфогенетических факторов (вместе с факторами внешней среды). Их удельный вес весьма высок в развитии половых признаков позвоночных животных и особенно птиц и млекопитающих. Однако в этом нет ничего принципиально отличного от гормональных влияний на развитие других организационных признаков (например, на развитие костного скелета).
В отличие, однако, от факторов индивидуального развития видовых (расовых) различий, факторы, обусловливающие развитие половых признаков известного вида организмов, обнаруживают лишь количественные различия между полами, а не качественные. Именно в этом заключается своеобразие механизма развития половых признаков. На почве разных количественных соотношений в генетических факторах возможно возникновение локальных качественных различий в морфогенетических процессах, которые могут в дальнейшем углубляться вследствие стимулирующих и тормозящих влияний со стороны системы половых гормонов, нa этой основе и возникают все новые половые различия (через интегрирующее действие естественного и полового отбора). Однако их поддержание на известном уровне у самца и у самки предполагает наличие устойчивости в определяющих их специфику генетических факторах и в концентрации стимулирующих и тормозящих гормонов. Концентрация половых гормонов всегда подвержена значительным колебаниям (индивидуальным уклонениям, сезонным изменениям и изменениям под влиянием всевозможных внешних факторов и общефизиологического состояния организма). Влияние этих колебаний в концентрации гормонов преодолевается развитием системы физиологических и морфогенетических регуляторных механизмов. Последние связаны с выработкой известных норм реактивности тканей, ограждаемых достаточно значительным промежутком между нижним и верхним порогами нормальной реактивности тканей. Изменение этой реактивности, т. е. положения пороговых уровней, достигается, однако, в свою очередь, изменениями в генетических факторах. При количественном характере генетических половых различий это требует всегда точной согласованности с другими факторами, определяющими развитие половых признаков и имеющими также чисто количественный характер (концентрация одних и тех же половых гормонов у самца и у самки). Поэтому стабилизация половых признаков через смещение пороговых уровней нормальной реактивности тканей у одного пола может привести к нарушению развития половых признаков другого пола. Между тем, основным механизмом стабилизации развития как раз и является смещение пороговых уровней, и именно — снижение нижнего порога и поднятие верхнего порога нормальной реактивности тканей (т. е. возможно широкое раздвигание обоих порогов).
При большой биологической роли и вместе с тем большой изменчивости (лабильности и мобильности) мужских половых признаков ведущим и в развитии половых различий, и в их стабилизации оказывается мужской пол.
Наличие процесса снижения нижнего порога нормальной реактивности тканей по отношению к стимулирующему мужскому гормону или «повышение чувствительности» тканей в процессе эволюции самцовых вторично-половых признаков птиц доказано во многих случаях (см. приведенные данные Б. Г. Новикова). Понятно, что по отношению к тормозящему влиянию овариаль-ного гормона нормальная мужская реактивность тканей должна изменяться в противоположном направлении. Эволюция самцовых признаков сопровождается снижением «чувствительности» тканей к женскому гормону (см. данные Б. Г. Новикова), т. е. повышением нижнего порога нормальной реактивности.
Стабилизация половых признаков самца является, очевидно, результатом действия стабилизирующей формы отбора. В данном случае стабилизирующий отбор покоится на элиминации самцов с недостаточным развитием половых признаков (вследствие высокого порога нормальной реактивности тканей при недостаточной концентрации мужского гормона или других неблагоприятных внешних и внутренних влияниях). Если в результате стабилизирующего отбора по самцам происходит снижение нижнего порога нормальной «самцовой» реактивности тканей (т. е. повышение их «чувствительности» к мужскому гормону), то при известной степени снижения он может достигнуть уровня нормальной концентрации мужского гормона у самки. В этом случае самцовая морфогенетическая реакция неизбежно реализуется и у самки.
Некоторые признаки развиваются у самцов как «независимые», а у самок не проявляются вследствие противодействия со стороны овариального гормона. Стабилизация таких признаков у самца идет (в силу той же элиминации самцов с недоразвитыми половыми признаками) по пути снижения «чувствительности» к тормозящему влиянию женского гормона, имеющегося нормально и у самца. При наличии колебаний в концентрации женского гормона у самца, эффективность стабилизирующего отбора может оказаться весьма значительной. На известном уровне стабилизации ткани могут утратить «чувствительность» к тормозящему влиянию даже тех концентраций овариального гормона, которые (встречаясь изредка у самцов) соответствуют норме для самки. Тогда «независимый» самцовый признак развивается и у самки.
Передача вторичных половых признаков на другой пол у ракообразных и у насекомых связана, очевидно, со стабилизацией реакций на генетические и морфогенетические факторы развития (которые в отношении половых признаков покоятся на количественных различиях). У позвоночных животных к ним прибавляются еще гормональные факторы, и в некоторых случаях их роль оказывается весьма значительной. Механизм стабилизации у одного пола и связанной с этим передачи на другой пол остается, однако, в общем одинаковым.
Механизмы развития, стабилизации и передачи первичных половых признаков также принципиально не отличаются от механизмов развития стабилизации и передачи вторичных половых признаков. Они развиваются, как правило, лишь на более ранних онтогенетических стадиях, когда роль гормонов еще не столь велика. Значение гормонов, однако, ясно выступает, например, в известных близнецовых интерсексуальных самках типа free-martin, наблюдаемых у рогатого скота в случаях установления связи в плацентарном кровообращении у разнополых близнецов (по Лилли). То же самое показано экспериментами Greenwood and Blyth (1930) с курами, когда в результате имплантации ткани семенника в курицу, у последней недоразвивались яйцеводы, а также не развивалась нормальная эластичность и широкие раздвигание костей таза. Все же несомненно доминирующее значение имеют в этом случае генетические и морфогенетические факторы развития.
Передача первичных половых признаков с одного пола на другой начинается всегда с эмбриональных зачатков. Это указывает на значение изменения генетических и морфогенетических факторов уже в первых фазах стабилизации этих признаков. Несомненно, это определяется их большим историческим прошлым, их значением и связанностью с общей организацией. С другой стороны, и вторичные половые признаки, по мере их стабилизации, развиваются на более ранних стадиях.
Еще Дарвин указывал, что передача полового признака на другой пол и превращение его в видовой признак сопровождается более ранним его онтогенетическим развитием.
Это более раннее развитие могло бы быть связано с более ранним вступлением в действие известных морфогенных факторов, определяющих развитие этих признаков. Оно могло бы быть связано с более ранней инкреторной функцией зачатка половой железы и ранним достижением порогового уровня концентрации половых гормонов (без изменения дефинитивной концентрации и порога нормальной реактивности тканей). Это не привело бы к передаче признака на другой пол, так как механизм развития половых признаков оставался бы ненарушенным. Однако общее повышение концентрации мужского гормона или уменьшение концентрации женского гормона у обоих полов (при сохранении нормального соотношения между этими концентрациями у разных полов) могло бы привести и к стабилизации самцовых признаков у самца и к их передаче на самку.
Изученные у птиц факты переноса самцовых признаков на самку показывают, однако, что этот перенос покоится в этих случаях на иных основаниях. Мы видим, что меняется не время поступления гормонов, не их концентрация и не их качество. Решающую роль играет изменение общей обоим полам нормальной реактивности тканей. Происходит снижение порога нормальной реактивности тканей на мужской гормон. Ткани реагируют на более низкие концентрации гормона. Понятно, что эта реакция становится тогда возможной и в более раннем возрасте.
Эволюция самцового признака, его стабилизация и более раннее развитие происходят непрерывно и постепенно. Однако в тот момент, когда нижний порог нормальной реактивности тканей на мужской гормон, постепенно снижаясь, достигнет наличного уровня его концентрации хотя бы у некоторых самок, сразу разовьются самцовые признаки. Таким образом, получается нередко наблюдаемая картина диморфных самок (например, безрогих и рогатых самок северного оленя). При дальнейшей стабилизации самцового признака у самца, через снижение порога нормальной реактивности тканей на мужской гормон, все самки приобретут данный самцовый признак. Это происходит «скачкообразно» в силу общих законов реагирования тканей, обладающих выдифференцированными «нормами», огражденными известными порогами (закон реагирования по типу «все или ничего».) Ближайшие соседние виды или даже подвиды и расы данного вида могут обладать нормальными самками.
У нас имеются все основания думать, что таков именно механизм передачи на другой пол не только тех признаков, которые обнаруживают явную зависимость от половых гормонов, но и таких признаков, которые, подобно первичным половым признакам, развиваются в известной степени независимо от гормональных влияний, по крайней мере на стадиях ранней закладки. При взаимосвязанности различных первичных половых признаков между собой (а у позвоночных и с выделительной системой), определяемых в своем развитии сложной системой факторов (корреляций), скачкообразный характер передачи до известной степени теряется. Передача начинается с ранних стадий развития и при непрерывном контроле со стороны естественного отбора (элиминация уклонений в половой системе) приводит к выработке некоторой общей для обоих полов основы, на которой еще возможна дальнейшая половая дифференцировка. Все же ясно выступает процесс передачи самцовых признаков (копулятивного органа) на самку. Иногда он достигает максимальной степени во внешней неотличимости самки от самца (пятнистая гиена). Ясно виден и противоположный процесс — стабилизация женских признаков и их передача на самца («мужская матка» бобра).
Само собой разумеется, что в случае ясно выраженной вредности самцового признака для самки начинается немедленно элиминация самок, обладающих самцовыми признаками. Таким образом, естественный отбор начнет противодействовать процессу превращения самцового признака в видовой и стабилизация первого остановится на известном уровне. То же самое относится и к стабилизации признаков у самок и их передаче на самцов.
Передача признака с самца на самку происходит в процессе эволюции на известном этапе его стабилизации. Этот процесс непосредственно не связан с прогрессивным развитием данного признака у самца. Но он, конечно, может быть связан с изменением его биологического значения.
Передача черной пигментации клюва воробьиных птиц с самца на самку, передача самцового оперения ушастых фазанов на самку, передача петушиной шпоры на самку некоторых куриных вовсе не связаны обязательно с их прогрессивным развитием у самцов. Эти процессы в значительной мере независимы. То же самое касается и передачи первичных половых признаков на другой пол. Развитие самцовых наружных органов у самки пятнистой гиены вовсе не связано с особым их развитием у самца данного вида, и развитие матки с яйцеводами у самца бобра также не определяется ее особым развитием у самки (она у нее ничем не отличается от матки самки канадского бобра, у которого «мужская матка» отсутствует).
Это указывает вновь на то, что передача признаков самца на самку не связана с явным изменением фенотипа самца, а следовательно (если она не связана с пользой для самки), не определяется обычной ведущей формой естественного (включая половой) отбора по самцовым признакам. Передача половых признаков связана здесь не с изменением дефинитивного фенотипа, а лишь со стабилизацией этих признаков у одного пола (обычно у самца). Этот процесс определяется стабилизирующей формой естественного отбора, идущего на основе селекционного преимущества установившейся нормы перед всеми от нее уклонениями. В данном случае основным объектом элиминации оказываются самцы с недостаточным или запоздалым развитием половых признаков (основанном на слабой чувствительности тканей к мужскому половому гормону), т. е. особи с недостаточно низким порогом нормальной «мужской» реактивности тканей. Это ведет к прогрессивному снижению нижнего порога реактивности и к вытекающим из этого, на известном этапе, последствиям для самки. Впрочем, к такому же результату привело бы и прогрессивное возрастание концентрации половых гормонов или других морфогенных факторов при сохранении их соотношений у обоих полов и сохранении общей для обоих полов реактивности тканей на прежнем уровне. И этот путь стабилизации признаков у одного пола мог бы приводить к их передаче на другой пол. Пока мы еще не имеем фактов, показывающих реальное осуществление такого пути стабилизации и передачи половых признаков.
Что в ходе эволюции действительно имеет место процесс стабилизации форм, видно из следующего.
1. Нередко локальные, экологические или географические формы весьма сходны с модификациями того же или близкого вида, однако отличаются от них тем, что сохраняют свои характерные признаки константными и при перенесении в условия, нормальные для этих близких форм или видов. Иными словами, локальное изменение оказывается наследственным, т. е. местная форма — более стабильной.
2. В некоторых случаях доказана замена в процессе эволюции внешних факторов развития внутренними, т. е. постепенная автономизация развития.
3. Многие функциональные структуры, которые впервые возникли под влиянием функции, развиваются автономно. Некоторые вторичноиоловые признаки возникают как признаки, зависящие в своем развитии от полового гормона, а затем становятся в процессе эволюции независимыми.
4. Установлен факт прогрессивного развития регуляторных механизмов онтогенеза в генетических системах, в системах градиентов и в индукционных системах, выражающийся в устойчивости и в доминировании нормы, при которых возможны известные сдвиги внутри взаимодействующей системы без нарушения нормального формообразования.
Что стабилизация форм достигается через посредство естественного отбора, доказывается целым рядом косвенных данных.
1. Трудно себе представить развитие регуляторных механизмов иначе, как только элиминацией вредных уклонений от нормы при случайных изменениях в факторах среды или при заметном выражении мутаций.
2. В случае «параллелизма» модификационных и наследственных изменений сходство достигается нередко разными путями, и, следовательно, нельзя говорить о простом «фиксировании» модификаций.
3. При переносе уклоняющейся экологической или географической формы в исходную среду наблюдается обычно лишь частичный возврат в некоторых только признаках к исходным формам. По меньшей мере в части признаков наступает наследственная стабилизация местной формы, что указывает на действие естественного отбора наследственных изменений в пределах модифицированного фенотипа.
4. Развитие ассимилирующей ткани в стеблях и черешках растений, сбрасывающих листья во время засухи, привело у некоторых ксерофитов к развитию своеобразных безлистных форм (или к развитию вторичных листообразных органов). Это показывает достаточно убедительно, что именно модификационные изменения в тканях стеблей и черешков были реальным этапом на пути эволюции этих форм. Однако дальнейшие наследственные изменения могли возникнуть лишь в результате естественного отбора.
5. Факты замены в процессе эволюции внешних факторов развития, вызвавших к жизни данную модификацию (воды, сухости), другими, дающими более постоянную и, следовательно, более надежную реакцию (влажностью, освещением), указывают уже на известную стабилизацию путем элиминации всех особей, давших «ошибочную» . реакцию. На то же самое указывают и факты замены внешних факторов развития внутренними и. функциональных факторов — морфогенетическими.
Усложнение системы морфогенетических корреляций, характеризующее автономное развитие и не сопровождающееся заметным изменением фенотипа, объяснимо также только действием стабилизирующего отбора мутаций, лежащих в пределах установившегося фенотипа (т. е. элиминацией всех заметных уклонений от «нормы»).
6. Прямыми доказательствами существования стабилизирующего отбора являются многочисленные наблюдения на лабораторных и полевых культурах, показывающие изменение генотипической структуры известных линий без заметного изменения их фенотипа. Генотипические изменения устанавливаются в этом случае лишь методами генетического анализа. То же самое показывает генетический анализ природных популяций дрозофилы, скрывающих под обычной внешностью нормального «дикого» типа всевозможные комбинации мутаций, различные в разных популяциях. Стабилизирующий отбор приводит и в лабораторных культурах к подавлению выражения вредных мутаций в гетерозиготе, т. е. к эволюции их рецессивности. В результате стабилизирующего отбора норма становится доминантной по отношению к мутациям и приобретает максимальную устойчивость как по отношению к изменениям во внешних факторах, так и по отношению к нарушениям во внутренних факторах развития.
Г. ЕДИНСТВО И ВЗАИМОДЕЙСТВИЕ ВЕДУЩЕЙ И СТАБИЛИЗИРУЮЩЕЙ ФОРМ ЕСТЕСТВЕННОГО ОТБОРА В ПРОЦЕССЕ ЭВОЛЮЦИИ
Современные представления о непрерывности процессов мутирования и перекомбинирования заставляют подойти по-новому и к пониманию естественного отбора. Последний выступает не только в роли движущего фактора, меняющего организацию соответственно изменениям в условиях существования. Даже при установившихся постоянных соотношениях между организмом и средой (с ее абиотическими и биотическими факторами), когда условия существования не меняются (за исключением периодических, временных или случайных изменений), проявляется непрерывное действие естественного отбора. Оно выражается по меньшей мере в постоянном противодействии разрушающему влиянию мутационной изменчивости и поддержании, через это, нормальной организации, приспособленной к данным условиям существования. Эта стабилизирующая роль естественного отбора проявляется решительно во всех условиях и в особенности обостряется в изменчивых условиях существования, когда общая изменчивость (как мутационная, так и модификационная) организмов заметно возрастает. Однако, когда условия существования начинают изменяться в определенном направлении и прежняя норма начинает терять свои преимущества перед некоторыми уклонениями от нее, естественный отбор приобретает роль фактора, ведущего к изменению установившейся организации.
Так как в историческом развитии организмов условия их существования непрерывно меняются, а в каждый данный момент они обладают известной определенностью, то ведущая форма естественного отбора всегда сопровождается стабилизирующей.
Изолированное рассмотрение ведущей и стабилизирующей форм естественного отбора представляет результат абстракции. Конкретно мы имеем всегда сосуществование обеих форм отбора. Точнее, мы имеем единый процесс естественного отбора с двумя его характерными выражениями.
Механизм естественного отбора и его результат в различных случаях нетрудно себе представить на обычных вариационных кривых. Постоянный процесс мутирования и постоянный процесс включения, комбинирования и размножения мутаций в каждой данной популяции должен был бы привести к общему увеличению изменчивости, т. е. к расширению вариационной кривой. Однако, вопреки процессам мутирования и распространения мутаций в популяциях, вариационная кривая сохраняет нередко свою форму неизменной или даже происходит ее сжатие. Это объясняется постоянным противодействием естественного отбора, идущего в пользу установившейся нормы (при элиминации всех значительных уклонений). Поэтому говорят не только о мутационном «давлении», но и ему противоположном «давлении» естественного отбора или, лучше сказать,— избирательной элиминации. Если первое определяет расширение вариационной кривой и снижение ее вершины, то второе определяет ее сжатие и поднятие вершины. В любых конкретных условиях существования устанавливается известное равновесие между обоими противоположными процессами и вариационная кривая приобретает определенную форму, типичную для известной популяции в данных условиях существования. Равновесие это имеет подвижной характер: увеличение изменчивости ведет к увеличению интенсивности элиминации (отбора), сокращающей индивидуальную изменчивость. Всякое улучшение условий, связанное с сокращением элиминации и, следовательно, с ослаблением интенсивности отбора, ведет поэтому к расширению вариационной кривой (действием мутационного «давления»). Однако то же улучшение условий ведет и к увеличению численности вида, обострению борьбы за существование, усилению избирательной элиминации, т. е. увеличению интенсивности отбора и, следовательно, к сужению вариационной кривой и крутому поднятию ее вершины (вследствие элиминационного «давления»). Если же крайние варианты одного направления получают известные преимущества в борьбе за существование перед крайними вариантами другого направления, то произойдет расширение вариационной кривой в первом направлении (в силу мутационного «давления») и сжатие ее на противоположном конце (под «давлением» элиминации). В результате совместного влияния мутирования и отбора будет происходить движение вариационной кривой в благоприятствуемом направлении. В этом движении обнаружится «ведущая» роль естественного отбора в процессе прогрессивной эволюции. В благоприятствуемом направлении будет происходить накопление наследственных изменений, ведущих к выработке новых приспособлений, а также новых форм приспособительного реагирования. В последнем случае можно говорить о «лабилизации» формообразования, приобретающего более зависимый характер.
В целом эволюция представляет сложный процесс непрерывной «лабилизации» признаков индивидуально-условного значения и «стабилизации» тех формообразовательных реакций и признаков, адаптивность которых приобрела постоянное значение. Эволюция — это непрерывный безостановочный процесс включения все новых вариантов, элиминации всех менее жизненных особей и непрерывный процесс создания более приспособленных и более стойких форм организации. Естественный отбор, как единый процесс, является ведущим фактором не только в непрерывном приспособлении организма к условиям его существования, а через это и в создании новых форм организации, но и в создании более или менее сложных и стабильных механизмов индивидуального развития. Сама эволюция приобретает характер не только направленного, но и довольно устойчивого движения.
7. ТВОРЧЕСКАЯ РОЛЬ ИНДИВИДУАЛЬНОГО ОТБОРА
В ведущей форме отбора наиболее ясно обнаруживается творческая роль естественного отбора, создающего новые формы. В стабилизирующем отборе лишь охраняется уже существующая норма. Она как будто не претерпевает видимых изменений. Если, однако, говорить о генотипе, о его норме реакций и о фенотипе в широком смысле, включая весь процесс осуществления известной нормы, то становится ясно, что и стабилизирующий отбор имеет весьма важное творческое значение.
Если он и не приводит непосредственно к созданию новых дефинитивных форм, то в результате его действия создается новый аппарат индивидуального развития, с его регуляторными механизмами. Происходит автономизация развития, связанная с последовательной заменой внешних факторов развития внутренними. Зависимые процессы развития преобразовываются D авторегулярные или они приобретают далее вполне независимый — автономный характер. Это означает изменение генотипа с его нормой реакций, так как реакция на внешний раздражитель заменяется подобной же реакцией на внутренний раздражитель. Реактивность тканей явно изменяется, а частично, очевидно, меняется и система формативных раздражителей. Во всяком случае, несомненно изменяется и генотип организма, приобретающего более устойчивые формы. В обоих случаях, и в ведущей форме отбора и в стабилизирующей его форме, естественный отбор имеет индивидуальный характер — он основан на элиминации менее приспособленных особей. Только этот индивидуальный отбор и имеет непосредственно творческое значение. Семейный или групповой отбор получает творческое значение лишь через индивидуальный отбор. В основном же групповая элиминация ведет лишь к вытеснению одной формы другой, более приспособленной в условиях данной местности. Правда, это ведет также и к очень важному результату — межгруповое соревнование в скорости приспособления ведет к переживанию более пластичных линий и к общему ускорению темпов эволюции. Любая форма естественного отбора связана с разрушением признаков (а следовательно — реакций и корреляции, утративших свое значение) и с созданием новых признаков, реакций и корреляций. Обычный естественный отбор связан соответственно как с изменением генотипа, так и с изменением фенотипа в направлении прогрессивного приспособления организма к условиям внешней среды. Стабилизирующий отбор также связан с разрушением известных реакций и корреляций, утративших свое значение. Фенотип зрелого организма при этом заметно не меняется. Но стабилизирующий отбор имеет и свою положительную, т. е. созидательную, роль. В процессе стабилизирующего отбора создаются новые морфогенетические корреляции, заменяющие в качестве внутренних факторов развития внешние факторы развития и морфогенетические реакции на эти факторы (автономизация развития). С созданием внутреннего формообразовательного аппарата, определяющего реализацию нормальной формообразовательной реакции вне обязательных рамок внешних факторов определенной интенсивности, развитие становится до известной степени автономным. Создание такого внутреннего аппарата развития, т. е. системы морфогенетических корреляций (в широком смысле), и является основным выражением творческой роли стабилизирующего отбора. Лишь после этой стабилизации форм, установившихся в результате приспособительной реакции, т. е. после создания внутреннего механизма их развития, можно говорить о действительно наследственном их закреплении.
В заключение отметим вкратце основные процессы, протекающие под направляющим влиянием естественного отбора.
1. Изменение плодовитости, как побочный результат общей элиминации, которая ведет к повышению плодовитости вследствие преобладающего переживания потомства более плодовитых линий. Повышение плодовитости, увеличивая численность особей, способствует увеличению абсолютного числа мутаций и их комбинаций. Общая элиминация ранних стадий (возрастная элиминация) ведет также к отбору особей с более быстрым развитием, а общая элиминация зрелой формы связана с отбором на раннее созревание и однократное размножение. Все это способствует быстрой смене поколений и, соответственно, ведет к ускорению эволюционного процесса, если элиминация приобретает индивидуальный характер. Обратное направление отбора, при косвенной индивидуальной элиминации, ведет к специализации, уменьшению плодовитости, к более позднему и длительному размножению и сокращению эволюционной пластичности (Шмальгаузен, 1939а).
2. В процессе отбора (при скрещиваниях внутри популяции) создаются под его контролем наиболее жизненные комбинации. В этом комбинировании мутаций меняется их выражение — благоприятные выражения усиливаются, а неблагоприятные выражения изменяются или полностью подавляются.
3. В процессе естественного отбора меняется и сама м у т а-бильность. Самое накопление мутаций ведет уже к повышению мутабильности. Кроме того, возможен и отбор мутабильных линий как побочный результат отбора прогрессивных форм, эво-люирующих с большей скоростью. Обратный процесс сокращения мутабильности является результатом стабилизирующего отбора, особенно в однородных и постоянных условиях внешней среды.
4. В результате естественного отбора на приспособленность к данным условиям среды меняется норма реакций организма и, в частности, устанавливаются наиболее выгодные формы реагирования на изменения в факторах внешней среды, т. е. создаются системы адаптивных реакций организма, наиболее сложные у лабильных организмов (лабилизация формообразования).
5. В процессе естественного отбора на максимальную организованность (согласованность функций, жизнеспособность, экономичность) устанавливаются' наиболее выгодные системы корреляций, обеспечивающих согласованное развитие частей и органов и их взаимную приспособленность (коадаптацию) в целостном организме. Эти корреляции охватывают не только формообразовательные процессы, но и биохимические процессы, лежащие в их основе и выражающиеся во взаимодействии ядра и плазмы (генетические корреляции).
6. В процессе стабилизирующего отбора (при элиминации всех уклонений от приспособленной нормы) создаются регуля-торные механизмы, обеспечивающие наиболее надежное развитие нормальной организации (или отдельных адаптивных норм) и при случайно уклоняющихся условиях внешней среды. В частности, система морфогенетических корреляций регуляторного характера, способствующая устойчивости формообразования, создается именно в процессе стабилизирующего отбора. Авторегуляторное и автономное развитие организмов представляет различные формы такой защиты нормального формообразования от нарушения со стороны случайных влияний факторов внешней среды. Однако защитные механизмы охватывают собственно весь аппарат индивидуального развития, как часть наследственного аппарата, и даже этот последний в целом (см. далее III—4). Наибольшей сложности достигает система морфогенетических корреляций, а следовательно — и устойчивость формообразования у стабильных организмов с регуляционным типом развития (именно у позвоночных животных).
8. ПАНМИКСИЯ И ИЗОЛЯЦИЯ
Благодаря непрерывной деятельности естественного отбора, любой вид организмов не только вполне приспособлен к той нормальной обстановке, в которой он обитает, не только обладает вполне согласованной, гармоничной организацией, но характеризуется и своими реакциями, приспособленными к обычным изменениям во внешней среде (целесообразным поведением у животных) и наличием тончайшего аппарата наследственности и развития. Особое внимание мы обращаем на существование сложной системы регуляторных механизмов, защищающих нормальное формообразование особи от возможных его нарушений со стороны различных как внешних, так и внутренних факторов. Неудивительно, что при этой сложной согласованности реакций организма любая мутация оказывается неблагоприятной. Она неблагоприятна даже тогда, когда она не проявляется, т. е. не имеет видимого выражения. Каждая мутация связана с изменением аппарата индивидуального развития и реактивности организма, и если сдвиг во времени или интенсивности раздражителей не превышает порогового уровня известной реакции и не нарушает ее нормального течения вследствие наличия регуля-торного механизма, то все же она всегда означает приближение к пороговому уровню реактивности. Поэтому любая мутация снижает защищенность известных формообразовательных реакций, делает их более доступными для других нарушающих влияний, т. е. по меньшей мере уменьшает устойчивость нормального формообразования (рис. 2, 3).

Рис. 15. Типичное строение крыльев и гальтеров всех возможных комбинаций аллеломорфов vestigial у Drosophila melanogaster. (No-wing, vestigial, noched, nicked и «дикая» норма), Цифрами обозначен процент краевых вырезок и относительный количественный объеме для каждой комбинации. По О. Моор, 1932

Рис. 16. Строение антенн различных аллеломорфов aristopedia и их комбинаций. Первый рисунок (+) показывает нормальную антенну «дикого» типа Drosophila тelanogaster. По К. Уоддингтону
Если отдельная мутация означает нарушение формообразовательного механизма, то простое суммирование их может вести только к его полному разрушению (рис. 15, 16).
Прежде всего — это само собой разумеется — бесконтрольное суммирование мутаций означает разрушение наследственного аппарата вместе с его защитными механизмами («генный баланс», доминирование нормы и т. и.). Во-вторых, это означает разрушение регуляторного механизма индивидуального развития, защищающего его нормальное течение от возможных нарушений (о чем только что было сказано). И, наконец, это означает разрушение вообще всего реакционного и корреляционного аппаратов, т. е. всей системы факторов индивидуального развития, ведущее к полной дезорганизации, к уничтожению приспособленности организма к внешней среде и к нарушению внутренней согласованности частей и функций. Такой распад организации наблюдается всегда в отношении тех органов и приспособлений, которые в процессе эволюции теряют свое значение. Редукция органов объясняется именно бесконтрольным накоплением мутаций (Шмальгаузен, 1938а). Если этого не происходит в процессе обычного мутирования любого вида организмов, то это объясняется постоянным действием естественного отбора, вводящего этот процесс в определенное русло.
А. СКОРОСТЬ ЕСТЕСТВЕННОГО ОТБОРА
Возможность естественного отбора определяется, конечно, наличием материала для отбора, т.. е. генетическим многообразием особей данной популяции. Максимально возможная скорость естественного отбора определяется масштабом изменчивости в признаках, имеющих какое-либо значение в борьбе за существование. В математическом выражении эта зависимость известна как основной закон селекции Р. Фишера (R. Fisher). По этому закону скорость отбора признака пропорциональна квадрату среднего квадратического уклонения данного признака, т. е. варианте. Изменчивость особей популяции зависит от ее насыщенности различными мутациями, которая бывает различной для разных мутаций в зависимости от частоты их возникновения и от биологического их значения. Большинство мутаций неблагоприятны для их обладателей, но некоторые из них могут давать частичные преимущества в некоторых локальных, сезонных или просто случайных условиях существования. Однако даже безусловно вредные мутации, если они рецессивны, могут широко распространиться в популяции. Во всех случаях распространение мутаций зависит от частоты повторного воьникновения данной мутации (мутабильности) и скорости ее уничтожения под влиянием естественного отбора (при полной рецессивности только после появления гомозиготных мутаций). Между мутационным «давлением» и «давлением» естественного отбора устанавливается некоторое равновесие на разном уровне концентрации для разных мутаций — более высоком для условно благоприятных или «индифферентных» мутаций и более низком для вредных и в особенности для полудоминантных деталей. Такое разнообразие в генетическом составе популяции и, особенно, большое число гетерозигот по разным мутациям составляют как бы основной фонд, или резерв наследственной изменчивости, служащий очень благоприятной основой для эффективного действия естественного отбора и быстрой перестройки генетической структуры популяции. По мере этого некоторые мутации элиминируются, другие, наоборот, суммируются, частью переходят в гомозиготное состояние и входят в состав новой нормы. Скорость естественного отбора зависит не только от изменчивости популяции, т. е. резерва уже накопленных изменений, но и от скорости мутирования. Возникновение неблагоприятных мутаций замедляет течение отбора, а возникновение благоприятных ускоряет его. Однако скорость мутирования вообще очень незначительна, да и отбору подлежат не отдельные мутации, а сложные их. комбинации в виде тех или иных фенотипов. Число таких комбинаций, конечно, неизмеримо больше числа мутаций и о недостатке материала для отбора, идущего всегда по фенотипам, не может быть и речи. Во всяком случае мутирование при определенной его скорости, хотя и может повлиять на общую скорость естественного отбора, не может изменить его направления и закономерного распределения скоростей в зависимости от величины коэффициента селекции и от концентрации отбираемых вариантов. Равномерное мутирование может привести лишь к вычитанию или прибавлению некоторой постоянной величины к приводимым далее значениям скоростей отбора.
Вопрос об эффективности естественного отбора теоретически разработан в целом ряде исследований Р. Фишера, Дж. Холдэна и С. Райта (R. Fisher, J. Haldane, S. Wright). Математический аппарат этих работ столь сложен, что не допускает доступного изложения. Мы отметим лишь немногие выводы. Прежде всего укажем, что по сделанным расчетам даже самые малые селекционные преимущества могут быстро привести к весьма заметным сдвигам в строении популяции. Большое значение имеют размеры популяции. В малых популяциях происходят родственные скрещивания, легко приводящие, при случайностях в элиминации, как к случайной утрате, так и к случайному фиксированию генов в гомозиготном состоянии. Эти случайные явления ускользают из-под контроля естественного отбора и, следовательно, естественный отбор в малых популяциях менее эффективен. Те же явления могут, однако, привести к возникновению новых комбинаций. В больших популяциях имеются гораздо более благоприятные условия для накопления самых разнообразных наследственных изменений. Случайные процессы практически никакой роли не играют, и возможности быстрого отбора очень велики. В больших популяциях, однако, весьма затруднено фиксирование благоприятных комбинаций. Очень большое значение имеют колебания численности популяции. В период максимального сокращения популяции родственные скрещивания ведут к гомозиготизации и фиксированию различных мутаций и их комбинаций в местах переживания остатков популяции. В период размножения естественный отбор ослаблен. Локальные популяции свободно размножаются, скрещиваются и дают начало множеству новых комбинаций, которые затем в период сокращения численности подвергаются самому жесткому отбору. Наиболее благоприятны для быстрого действия естественного отбора именно такие популяции с колеблющейся численностью или малые лишь частично изолированные популяции, которые систематически обмениваются мигрантами (...)
Кривая, выражающая изменение скорости отбора в зависимости от концентрации отбираемого варианта, имеет при малом коэффициенте селекции почти симметричную форму и, следовательно, наибольшая скорость отбора наблюдается на средних его концентрациях. При увеличении коэффициента селекции максимум скорости отбора сдвигается в сторону меньших концентраций и кривая распределения скоростей отбора приобретает асимметричную форму.
У диплоидного организма соотношения становятся более сложными, так как появляется третий, промежуточный тип, который может иметь различное селекционное значение. Соотношение трех типов в равновесной популяции определяется формулой Вейнберга — Гарди: (р + q)2 = р2 + 2pq + q2 = 1. В этом случае при простых мутациях имеются три генотипа АА, Аа и аа, которым соответствуют и фенотипы в числовом соотношении, указанном формулой. При определении скорости отбора, который идет всегда по фенотипам, приходится учитывать не только селекционное преимущество отбираемой гомозиготы, но и гетерозиготы (если она имеет хоть какое-либо отличие от элиминируемого варианта). Это усложняет расчеты, в которых теперь участвуют уже два коэффициента селекции. Ввиду обычного доминирования нормы мы можем рассмотреть лишь этот частный случай, что значительно упростит нашу задачу.
Скорость отбора находится в той же зависимости от селекционного коэффициента и от концентрации отбираемого варианта (фенотипа), как и в случае гаплоидов. Однако сама концентрация вариантов меняется иначе. В данном частном случае, при полном доминировании, гетерозиготы сходны с доминантными гомозиготами и это значительно увеличивает концентрацию доминантного фенотипа. Это ведет к ускорению отбора доминанты, особенно на малых концентрациях гомозиготы. Наоборот, концентрация рецессивного фенотипа оказывается уменьшенной, так как он в генотипе Аа не получает выражения и не может быть предметом отбора.
Таким образом кривые, выражающие распределение скоростей отбора фенотипов в зависимости от концентрации соответствующих генов, получают для доминанты и для рецессива различную форму. В особенности интересен, однако, частный случай отбора при наличии селекционного преимущества гетерозиготы перед обоими гомозиготами. При малом селекционном преимуществе гетерозиготы кривая распределения скоростей отличается от соответствующих кривых для гомозигот лишь большей растянутостью максимума, лежащего на средних концентрациях. При более остром отборе кривая приобретает однако ясно седлообразную форму, т. е. двухвершинный характер. В случае равного преимущества гетерозиготы перед обеими гомозиготами эта кривая оказывается вполне симметричной.
Приведенные формулы, а также кривые распределения скоростей отбора в зависимости от концентрации отбираемого гена в популяции позволяют сделать некоторые интересные заключения. Скорость естественного отбора зависит как от концентрации отбираемого варианта (р), так и от концентрации элиминируемого варианта (д) или, иными словами, она пропорциональна интенсивности соревнования, которую мы измеряем произведением обеих концентраций (w = apq). Скорость отбора зависит также от селекционного преимущества одного варианта перед другим. При малом коэффициенте селекции скорость отбора просто пропорциональна этой величине. При более значительных различиях в конкурентоспособности зависимость сложнее и выражается в нашей формуле величиной s/(1-qs).
Если рассматривать скорость отбора в зависимости от концентрации (р) отбираемого фенотипа, то ясно, что перед нами прямая пропорциональность Δp=p(qs/(1-qs)). В этом случае можно сказать, что скорость отбора определяется не только концентрацией отбираемого варианта, но и его оценкой, которая дается в нашей формуле величиной qs/(1-qs). Оценка отбираемого варианта зависит как от величины коэффициента селекции, так и от концентрации элиминируемого варианта и достигает максимума при его максимальной концентрации. Это собственно понятно, так как оценка в борьбе за существование имеет сравнительный характер. Вариант А может выявить свое преимущество перед В лишь при совместном существовании обоих вариантов в популяции. Если вариант А редок, то его преимущества перед В сказываются с наибольшей ясностью. Если вариант А стал обычным, то он уже теряет свое выигрышное положение в популяции, в которой особи А преобладают. Если же вариант А завоевал всю популяцию, то отдельная особь А уже не имеет никаких преимуществ перед другими особями этой популяции. Другими словами, качество варианта не имеет абсолютного значения. Оно всегда релятивно. Оценка варианта производится всегда на фоне его альтернативы и является максимальной при решительном преобладании в популяции элиминируемого варианта. Чем значительнее коэффициент селекции (его пределом является s = l), тем выше оценка отбираемого варианта уже на самых малых его концентрациях. Поэтому уровень максимальной скорости отбора сдвигается при возрастании коэффициента селекции со средних концентраций при s>0,l все более в сторону меньших концентраций отбираемого варианта. Кривая распределения скоростей отбора в зависимости от концентрации отбираемого гена приобретает при этом все более асимметричную форму. В пределе, при 5=1, скорость селекции определяется только концентрацией элиминируемого варианта и, следовательно, максимальна уже на самых минимальных концентрациях отбираемого варианта (Ар = q).
Седлообразная форма кривой распределения скоростей отбора гетерозиготного фенотипа в зависимости от концентрации одного из генов является добавочной иллюстрацией сказанного. Кривая эта имеет два максимума — один максимум определяется оценкой гетерозиготы А а на фоне полного преобладания в популяции одной гомозиготы (аа) или одного гена (а), а второй максимум определяется оценкой той же гетерозиготы на фоне полного преобладания другой гомозиготы (АА) или гена А. Чем выше значение коэффициента селекции, тем шире раздвинуты оба максимума и тем яснее они выражены.
Все это имеет глубокий биологический смысл. Естественный отбор есть результат внутривидового соревнования особей (вариантов). Если скорость естественного отбора определяется при прочих равных условиях произведением из концентраций обоих вариантов, то это значит, что она пропорциональна напряженности их соревнования. Скорость естественного отбора пропорциональна, следовательно, математической вероятности столкновения обоих альтернативных вариантов. В начале отбора, когда концентрация отбираемого варианта А очень мала, каждая особь А сталкивается практически только с особями В и при этом полностью выявляются ее преимущества. Однако скорость естественного отбора мала, так как число особей А (а следовательно и общее число гетерогенных столкновений) еще очень мало. По мере нарастания концентрации особи А все чаще сталкиваются и с другими особями А и в этом случае не выявляют своих преимуществ. Однако скорость отбора нарастает, так как число особей А, а вместе с тем и абсолютное число столкновений между разными вариантами продолжают нарастать. Когда концентрация А достигает 50%, особи А сталкиваются в своей жизни в равном числе случаев как с особями В, так и с особями А. Только в первом случае выявляется их преимущество. Особи А утратили, следовательно, свое исходное преимущество в популяции по меньшей мере на 50%. Однако абсолютное число столкновений теперь максимально. При преобладании в популяции варианта А он сталкивается уже главным образом с особями А и, следовательно, окончательно лишается своих былых преимуществ в популяции. С падением концентрации варианта В уменьшается теперь и число гетерогенных столкновений и скорость отбора быстро падает.
При отборе гетерозиготы Аа максимум скорости отбора определяется выявлением ее преимуществ при наибольшей вероятности столкновения с гомозиготой аа в случае ее преобладания в популяции и вероятностью столкновения с гомозиготой АА при преобладании последней.
Таким образом естественный отбор определяется в своей скорости внутривидовым соревнованием вариантов, интенсивность которого зависит от распределения концентраций, т. е. от вероятности столкновения альтернативных форм между собой. Кроме этого, скорость естественного отбора определяется, конечно, селекционным преимуществом, выявляемым при сопоставлении этих вариантов в условиях данного биогеоценоза.
Кривая распределения скоростей отбора гетерозиготы замечательна не только по своей форме. Она демонстрирует большую эффективность естественного отбора гетерозиготного фенотипа в огромном диапазоне концентраций каждого из генов (от 0,1 до 0,9) или каждого из гомозиготных фенов (от 0,01 до 0,8).
Мы говорили пока только о скорости отбора фенотипов. Однако отбор фенотипов по необходимости связан и с отбором генотипов.
При отборе гаплоидов отбор фенотипа равнозначен отбору генотипа. В случае диплоидного организма это не совсем так. При полном доминировании скорость отбора доминантного гена снижается вследствие отбора гетерозиготных особей Аа, которые содержат в себе, кроме отбираемого гена А, также элиминируемый ген а. Снижается также скорость отбора рецессивного гена а, который отбирается только по относительно редкому фену аа и оказывается недоступным для отбора по обычным фенам Аа, Для отбора доминантных генов это уменьшение скорости отбора особенно значительно (примерно вдвое) вначале, когда почти все гены А заключаются в гетерозиготах. Однако по мере увеличения количества гомозиготных доминантов отбора гена А быстро возрастает, приближаясь к скорости отбора фена (АА + Аа), так как относительное число гетерозигот в популяции уменьшается. Наоборот, при отборе рецессивного фенотипа аа, отбор гена а идет вначале почти с той же скоростью, как и отбор фена аа, так как последний содержит только гены а. Однако по мере увеличения концентрации а, элиминируемые гены А сохраняются главным образом в гетерозиготах Аа, в которых гены а недоступны для отбора. Элиминация гетерозигот замедляет отбор генов а примерно вдвое (по сравнению со скоростью отбора фенотипа аа).
Для непосредственного определения изменения концентрации генов в популяции (за одно поколение) в результате естественного отбора диплоидных особей по их фенотипам была предложена следующая приближенная формула (Wright): Ар ~ pqs [q -r-+ hs (1–2q)], где р означает концентрацию доминантного гена А, q — концентрацию рецессивного гена a, s — селекционное преимущество фенотипа АА и hs — селекционное преимущество гетерозиготного фенотипа Аа перед гомозиготой аа. Эта формула пригодна только при малом значении селекционных коэффициентов s и hs и не малых концентрациях р (А).
При равенстве селекционного преимущества гетерозиготы и нормальной гомозиготы АА и, в частности, в случае полного доминирования нормы, вполне точные результаты дает формула Δp=pq2(s/1-q2s)), которая при малых значениях s дает (как и первая формула) приближенно: Δp≈pq2s.
Во всех наших рассуждениях мы считали, что коэффициент селекции является величиной постоянной для данной пары альтернативных вариантов. Это вряд ли так. При отборе положительного варианта А не только нарастает его концентрация, но при постоянном перекомбинировании могут улучшаться и его качества, в результате чего значение коэффициента селекции будет нарастать и это может привести к заметному ускорению отбора. Вероятно, такое ускорение будет наибольшим именно в первой фазе завоевания своего места в системе данной популяции.
Мы видели, что скорость отбора доминантного фена (и гена) возрастает при увеличении концентрации соответствующего гена в популяции вначале очень быстро. Наоборот, скорость естественного отбора рецессивного фена (и гена) нарастает вначале, при малых концентрациях, очень медленно, хотя и достигает позднее того же уровня, как и в случае доминантности. Это ведет при отборе в течение ряда поколений к очень большому отставанию в достижении заметного эффекта при отборе по рецессивным признакам. Однако конечный результат оказывается через некоторое время тем же самым.
Как бы ни было велико значение подобных теоретических расчетов, следует всегда помнить, что их правильность зависит от правильности принятых предпосылок и их значение поэтому всегда условно. Всегда в основу расчетов кладутся результаты известной схематизации, упрощения реальных соотношений. В действительности, например, не существует ни абсолютного доминирования, ни полной рецессивности. Следовательно, реальные кривые скорости отбора займут промежуточное положение между нашими теоретическими кривыми. Совершенно новые мутации не бывают рецессивными и если их выражение хотя бы в какой-либо мере благоприятно, то одновременно с их отбором начнется и отбор на усиление этого выражения в гетерозиготе. Мутация начнет доминировать и это быстро приведет к ускорению отбора, не только вследствие самой доминантности, но и вследствие возрастания коэффициента селекции. Кроме того, в природных условиях жизни амфимиктических популяций эволюция редко строится непосредственно на отборе вновь возникающих отдельных благоприятных мутаций. Обычно популяции находятся в состоянии некоторого генетического равновесия и при изменении условий существования переходят из одного стационарного состояния в другое за счет мобилизации внутренних резервов изменчивости. В этом случае все преимущества будут на стороне рецессивных генов, которые уже обладают, в отличие от доминантных, значительной концентрацией в любой природной популяции. При эволюционной перестройке популяции доминантные и рецессивные мутации стартуют на совершенно разных уровнях концентрации. Поэтому отбор рецессивных мутаций может произойти со значительно большей скоростью, чем отбор доминантных.
В случае выявления каких-либо преимуществ у гетерозиготы ее отбор произойдет именно в начале с исключительно большой скоростью, так как все мутации находятся в генетическом резерве равновесной популяции именно в гетерозиготном состоянии и нередко достигают высокой концентрации. Однако при свободных скрещиваниях и отсутствии специальных генетических механизмов, поддерживающих постоянную гетерозиготность, полное насыщение популяции гетерозиготной мутацией оказывается недостижимым (максимальный уровень насыщения достигается только при малой жизнеспособности гомозиготной мутации).
Б. КОМБИНИРОВАНИЕ МУТАЦИЙ
При половом размножении происходит не только распространение мутаций в потомстве, а следовательно — и в популяции, но и их разнообразное комбинирование. При свободном скрещивании (панмиксии) особей, обладающих генетическими различиями, создаются новые комбинации, которые в потомстве расщепляются и затем опять комбинируются, создавая при каждом скрещивании все новые и новые комбинации (рис. 15, 16). Если бы все комбинации были жизнеспособны и равноценны, то их возможное число достигло бы величин, поражающих воображение. Если бы генотип организма содержал лишь 1000 разных генов (несомненно, что у дрозофилы их больше) и каждый ген был способен к десяти изменениям (т. е. мог бы дать начало десяти аллеломорфам), то число возможных комбинаций было бы равно 101000, т. е. выражалось бы сверхастрономическим числом. Эти числа, конечно, не могут нигде реализоваться. Огромное большинство комбинаций было бы абсолютно нежизненным. Ничтожная доля возможных комбинаций все еще невероятно разнообразна и может быть представлена всеми переходами от полной летальности до высшего уровня приспособленности, достигнутой данным организмом. Можно эти реально возможные комбинации представить себе, по Райту, распределенными по полю, на котором низменности и «долины» означают низшие уровни жизнеспособности, а возвышенности с их «вершинами» соответствуют высшим уровням адаптации организма. В больших нерасчлененных популяциях имеется одна такая главная возвышенность, которая в случае падающей мутабильности и возрастающей интенсивности отбора сокращает свою площадь и становится более крутой, поднимая свою вершину до максимального уровня приспособленности. Такое состояние соответствует специализации.
При этих условиях возможность использования новых мутаций, которые лишь редко попадают в область главной возвышенности, очень низка, и такие виды могут при изменении условий оказаться недостаточно пластичными. Изменение факторов среды ведет к изменению «рельефа» площади, занимаемой комбинациями генов, и к постепенному перемещению вершины приспособленности в известном направлении, соответственно новым условиям существования.
Сокращение величины популяции ведет к уменьшению числа возможных комбинаций, к уменьшению изменчивости, к «фиксации генов» более или менее независимо от их адаптивной ценности. Поле комбинаций сокращается; «вершина» уменьшается и блуждает без определенного направления. Это — путь вымирания. Наконец, при разбивке вида на различные формы и местные расы, которые обладают своими более или менее обособленными вершинами приспособленности, судьба отдельных возвышенностей может быть весьма различной: одни из них сокращаются и исчезают, другие возникают вновь на месте прежних «долин», третьи перемещаются и расширяются или достигают высшего уровня специализации. Однако для вида в целом создаются, благодаря комбинации внутри группового и межгруппового отборов, наиболее благоприятные условия для прогрессивной эволюции, в особенности если отдельные популяции не вполне изолированы и постоянно обмениваются мигрантами.
Известный баланс между факторами эволюции («не слишком высокая мутабильность, не слишком жесткий отбор, не слишком тесный инбридинг и не слишком широкое скрещивание») является основным условием пластичности организмов (С. Райт).
В. ОГРАНИЧЕНИЕ ПАНМИКСИИ
Изоляция (постоянное ограничение панмиксии). Территориальное или биологическое ограничение скрещивания, раз установившись, сохраняет свое значение длительно и обычно со временем все более углубляется. Начинаясь как частичное ограничение свободы скрещивания, нарушаемое постоянной миграцией отдельных особей, оно дополняется, по мере установления различий между обособленными популяциями, постепенным развитием более существенных преград, рано или поздно полностью устраняющих возможность скрещивания. Наиболее общее значение имеет бесспорно дарвиновский фактор — элиминация менее приспособленных особей, устраняемых тем самым от возможности скрещивания с преуспевающими особями, которые в данных условиях оказались более (приспособленными. Из других возможных форм изоляции наиболее общий характер имеет, по-видимому, территориально-географическое расчленение, особенно существенное на начальных этапах обособления новых форм. На дальнейших этапах расхождения оно переходит в различные формы биологической изоляции или дополняется ими. Из этих последних следует учитывать значение экологической изоляции, не всегда отделимой от географической, значение различных форм физиологической (в том числе — половой) изоляции и, наконец, значение генетической изоляции, которая, проявляясь в стерильности гибридов, иногда устанавливается сразу, но чаще всего завершает процесс полного обособления новой формы от исходной (т. е. процесс видообразования).
Мы здесь рассмотрим значение раличных форм изоляции лишь как факторов, способствующих начальному сохранению и распространению новых комбинаций через ограничение их скрещивания с исходными формами, оставив в стороне большой вопрос о значении изоляции для видообразования и для дальнейшего расхождения органических форм.
1. Территориальная изоляция. Каково бы ни было значение географических преград, в виде непроходимых для известного вида пространств суши или воды, горных хребтов, пустынь, рек и т. п., мы не имеем в виду разбирать здесь эти вопросы, как общеизвестные и имеющие вместе с тем лишь частное значение в видообразовании. Мы остановимся на менее заметном и менее полном территориальном разграничении, которое имеет, однако, более общее значение, в особенности в начальных процессах дифференциации отдельных популяций в пределах вида или подвида. Вместе с тем как частичное, допускающее довольно широкий обмен особями между отдельными популяциями, оно имеет в большей мере прогрессивное значение, способствуя сохранению пластичности и возможности более быстрых темпов эволюции.
Такое частичное ограничение скрещивания достигается уже наличием самого факта размещения отдельных особей в пространстве, в особенности у малоподвижных или оседлых (хотя бы лишь в период размножения) животных. Его значение наиболее ясно у сидячих животных и у растений.
Сошлемся прежде всего на образование целых рядов форм в тех случаях, когда известный род или вид занимает большой ареал распространения и лишен возможности свободного перемещения в пределах этого ареала. В особенности это касается многих растений. Хорошим примером может служить сон-прострел (Anemone Pulsatilla), который широко распространен по континенту Евразии и дает непрерывный ряд форм, закономерно изменяющихся в направлении с запада на восток Европы. На западе подвид Л. p. ssp. germanica обладает тонко рассеченными и разбросанными листьями и поникшими цветами. На востоке у подвида А. р. ssp. grandis более грубо рассеченные, стоячие листья и стоячие цветы. Этот комплекс изменений связан с обилием дождей на западе и засушливостью восточной Европы (Zimmermann, 1938). Положение листьев способствует стеканию дождевой воды к корневой системе у восточных форм и, наоборот,— разбрызгиванию воды — у западных. Поникшее положение цветов предохраняет их от смывания пыльцы дождевой водой. Степень рассеченности листьев связана с различной интенсивностью транспирации. Мало рассеченные листья восточной формы испаряют заметно меньше воды, чем сильно рассеченные листья западной формы (что доказано измерением транспирации в одинаковых условиях опыта) . Приспособительность различий совершенно ясна, но вместе с тем — местные расы образуют еще совершенно непрерывный ряд форм в пределах вида между обоими крайними подвидами. Различия оказываются наследственными и типично расщепляются в потомстве гибридов. Однако одновременно имеются и модифика-ционные изменения, которые наслаиваются на наследственные.
Рыба бельдюга (Zoarces viviparus) образует в Скандинавии, по Шмидту, в отдельных фиордах совершенно непрывные ряды форм. В направлении от устья к концу фиорда у этой рыбы постепенно уменьшается число позвонков и число лучей некоторых плавников.
Африканские сомы рода Clarias образуют непрерывный ряд видов, ведущий от Clarias lazera Си v. et Val. с широкой лентой сошниковых зубов в Сев. Африке через С. mossambicus (Peters) из области озер Виктория и Танганьика и С. gariepinus (Burchell), распространенной от Замбези до Оранжевой реки, и до С. сареп-sis С. u v. et V а 1. с узкой прерванной полосой сошниковых зубов, живущей только в Натале (Worthington, 1940). Однако естественно, что на этих животных наиболее ясно видны результаты территориального разграничения у сидячих и малоподвижных форм. Поэтому классические примеры территориальной дифференциации относятся к наземным моллюскам (исследования Гулика, 1905, над улитками рода Achatinella на Гавайских островах, исследования Крэмптона, 1919, 1925, 1933, над улитками рода Partula на островах Таити и Муреа, исследования Плате, 1907, над улитками рода Cerion на Багамских островах), а частью и к донным, прибрежным формам (Purpura lapillus у Британских островов, по Куку). У морских моллюсков малая подвижность большинства форм компенсируется некоторой подвижностью личинок (в планктоне). Однако и планктонные организмы не рассеиваются особенно широко, даже когда они переносятся широкими массами в морских течениях, так как сохраняют пространственную связь между родственными особями. Даже активно плавающие и далеко мигрирующие организмы нередко держатся совместно, плавая целыми стайками. Образование многих элементарных экологических рас или линий у обыкновенной сельди (по Гейнке, Heincke, 1898) объясняется именно их инстинктами, благодаря которым целые выводки держатся в своих миграциях совместно и возвращаются для икрометания на прежние места. Это и поддерживает у них родственное скрещивание, дающее возможность закрепления вновь возникающих удачных комбинаций. То же самое касается, по-видимому, и различных лососевых и ряда других мигрирующих рыб. Тем более это относится к рыбам с оседлым образом жизни, подобным береговой бельдюге (Zoarces viviparus), которая образует множество местных рас, живущих в отдельных фиордах Скандинавии (Schmidt, 1917). Однако то же самое встречается и у таких подвижных животных, как птицы, в том числе перелетные. Обыкновенный аист (Ciconia ciconia), гнездящийся в Европе и зимующий в Африке, возвращается ежегодно IK одному и тому же гнезду, а молодь возвращается к местам своего вывода и гнездится в ближайшем соседстве. Фактически скрещивание происходит в очень тесной популяции родственных форм и лишь изредка освежается случайным прибытием мигранта.
То же самое касается ласточек. Меченая молодь возвращалась после зимовки на места своего рождения и гнездилась в пределах двух километров от материнского гнезда (Wachs, 1926). Ренш (Rensch, 1939) сопоставил результаты видовой дифференциации у больших птиц с широким распространением и миграцией, у малых перелетных птиц и у малых оседлых (по сводке птиц Пале-арктики Хартера). Из этого сопоставления ясно видно значение оседлости для дифференциации видов. Очевидно, родственное скрещивание (т. е. ограничение панмиксии) способствует выделению новых форм.
Интересный пример значения оседлости для самого начального разграничения популяций дали наблюдения Промптова над зябликом, который в южных областях СССР распадается на ряд популяций, ограниченных известным ареалом обитания и различающихся только по пению. Весьма вероятно, что эти различия не имеют еще ясно выраженной наследственной базы — молодь учится петь у родителей и других взрослых птиц ближайшего соседства. Такое обособление возможно, однако, лишь при условии возвращения после перелета в прежние места обитания (Промптов, 1930). В Московской области зяблик разбился на две экологические формы, различающиеся только по выбору мест для гнездостроения: одна гнездится в лесу таежного типа, смешанном со значительной примесью ели, а другая — в невысоких и редких, с полянками, молодых еловых и ??? рощах (Промптов, 1934)

Если это касается таких подвижных животных, как рыбы, с одной стороны, и птицы — с другой, то в еще большей мере то же применимо и к любым другим животным. Экспериментальные исследования Нобля (Noble, 1934) над ящерицей Sceleropus undulatus показали это с полной ясностью. Этот автор следил за переменой мест обитания ящериц, помеченных летом на их естественном местопребывании. Из 226 меченых ящериц к следующему лету сохранилось на прежнем месте 155. Многих из них видели многократно. Наибольшее отдаление местонахождений от исходного места достигало лишь 250 футов. При искусственном перенесении на более значительное расстояние ящерицы возвращались назад.
У автора создалось впечатление, что каждая особь охотится в пределах своей, очень ограниченной территории, лишь изредка заходя на территорию соседа.
Как известно, то же самое касается многих хищных птиц, а также и млекопитающих, держащих под своим контролем определенную территорию, необходимую для их прокормления. В особенности это относится ко многим грызунам, в том числе и крупным (бобры), а также к крупным хищникам.
Еще большее значение, чем обычная оседлость, имеет, конечно, сидячий образ жизни. Из животных он свойствен исключительно водным, так как только в воде может быть обеспечен пассивный перенос пищевого материала. С другой стороны, для растений только в воде возможна известная подвижность молодого или зрелого организма.
Прикрепленный образ жизни ведет, в общем, к тем же результатам, как и оседлость. Во всех этих случаях имеются средства для перемещения и распространения. Сидячие животные обладают подвижными личинками, а у растений пыльца переносится ветром или насекомыми и семена рассыпаются и подхватываются также ветром или высшими животными. Таким образом осуществляется панмиксия, а следовательно, и непрерывное перекомбинирование наследственного материала. Однако практически эта панмиксия значительно ограничена прикрепленным образом жизни. Подвижность личинок сидячих животных невелика, и они в большинстве прикрепляются поэтому недалеко от родительских особей, так что и скрещивание фактически ограничивается главным образом близкими родственниками. Да кроме того, сидячие животные часто обладают еще способностью к вегетативному размножению, т. е. к образованию генетически вполне однородных колоний. У растений, хотя пыльца и разносится, все же наиболее велики шансы опыления цветов собственной пыльцой, пыльцой с других цветов той же особи или от соседней особи. Семена же практически высыпаются в ближайшем окружении родительской особи. Образование родственных колоний поддерживается нередко способностью к вегетативному размножению.
Наконец, к тому же ведет и то обстоятельство, что почти никогда ни растение, ни животное не встречает необходимых для него экологических условий на больших сплошных пространствах. Обычно, даже при сплошном географическом распределении, подходящие экологические условия (почвы, их влажность, освещение, сообщество других организмов) встречаются лишь пятнами, которые тогда и оказываются средоточием колонии близко родственных особей. Величина таких колоний иногда ничтожна и при условии полной изоляции они были бы обречены на гибель, в силу утраты пластичности. Так как, однако, обмен мигрантами все же время от времени осуществляется, то и генетический состав популяций оказывается подвижным. Вместе с тем в каждой популяции фиксируются свои особенности, свои комбинации, которые испытываются в борьбе за существование и могут быть исходными для более широкого распространения.
В самом деле, как показали исследования Андерсона (Е. Anderson, 1936) над колониями Iris virginica, такие генетические различия между отдельными колониями вполне уловимы.
2. Экологическая изоляция. Экологическая изоляция, как упомянуто, не всегда отграничима от территориальной и географической, поскольку географические различия могут сопровождаться экологическими, а экологические формы разделяются и территориально. В особенности это касается растений, у которых, однако, иногда особенно ясно выступает именно экологическое расчленение как начальный этап образования новых форм. Таково значение почвенных условий и условий освещения в формировании экологических рас (экотипов), которые, возникая как экологические формы, оказываются сразу расчлененными и территориально, что способствует родственному скрещиванию внутри каждой новой группировки. Поскольку вегетация и время созревания у растений весьма зависимы от экологических условий, такое обособление может непосредственно привести и к различиям во времени созревания, и тогда экологическая изоляция дополняется биологическим ограничением возможности скрещивания с исходной формой. Так, по Андерсону (Е. Anderson), обособление близких видов Iris обусловлено в значительной мере расхождением во времени цветения. У животных некоторое расхождение в местообитании, в пищевом режиме, в образе жизни (дневной или ночной) связано с уменьшением шансов на встречу различающихся в этих отношениях особей, а следовательно, также способствует скрещиванию внутри обособливающихся групп. В основе этого расхождения лежит у животных, очевидно, некоторое изменение инстинктов, ведущее к изменению образа жизни, питания и т. п. Поэтому зоологи рассматривают иногда экологическую изоляцию как частную форму физиологической. Типичным примером начального обособления экотипов у животных могут служить песчаная и солончаковая формы муравья Cardiocondyla stambulovi For. (К. В. Арнольди, 1939).

Рис. 17. Некоторые байкальские бокоплавы. 1 — Brandtia lata lata (Dyb.); 2— Parapallasea lagowshii (Dyb.); 2— Ommatogammarus albinus (Dyb.); 4 — Garjajcwia sarsi (Sow.); 5 — Marcrochestopus branichii (Dyb.); 6 —Gammarasphaera insularis (Bas.); 7 — Acanthogammarus victorii (Dyb.); 8 — Spinacanthus parasiticus (Dyb.). По А. Я. Базикаловой, 1940
Типично экологический характер имеет процесс обособления нескольких рас мышей Peromyscus maniculatus, живущих в одном и том же ареале, но предпочитающих в одном случае лес, в другом случае — берега озер (Dice, 1931) или открытые места. Различия в окраске некоторых видов грызунов Perognathus связаны с различиями в цвете почв их местообитаний (Benson, 1933). Эти различия имеют экологический характер. Они связаны, однако, и с пространственным разграничением. Экологический характер имеет и распределение горных растений и животных на разной высоте. Множество эндемичных видов бокоплавов (gammaridae) оз. Байкал должно было образоваться на месте, на базе экологического расхождения немногих родоначальных форм по различным грунтам, зарослям, по разным глубинам и т. п. (рис. 17). Сезонное разделение произошло у рас обыкновенной сельди, которые мечут икру либо весной в различных местах прибрежного мелководья, либо осенью на мелях более открытого моря. Сезонные расы, отличающиеся по месту и времени икрометания, известны и для целого ряда проходных и полупроходных пресноводных рыб из семейств лососевых, осетровых, карповых, а также у судака (Л. Берг, 1934). Обыкновенная креветка (Crangon vulgaris) известна в двух расах, еще не различимых по виду, из которых одна мечет икру весной, другая — осенью (L. Plate, 1908). Дубовый шелкопряд (Lasiocampa quercus) появляется в июле — августе, а его почти неотличимый подвид (L. guercus callunae) — в мае — июне, так что скрещивание между ними почти невозможно. Бабочки Agriodes thetis и A. coridon различаются тем, что второй вид имеет один период размножения, падающий между двумя периодами размножения первого. Близкие виды бабочек — Eupithecia innotata Hufn. (выводится на Artemisia) и Е. unedo-nata Mob., которая выводится много раньше (на Arbutus), нормально не могут скрещиваться. Однако путем охлаждения куколок второго вида можно задержать вылупление, и тогда удается произвести скрещивание и получить вполне плодовитые гибриды Dietze. Такие же различия имеются между расами многих других бабочек (многие шелкопряды, в том числе тутовый), образующих различное число поколений в течение одного сезона. Фактически не скрещиваются в силу расхождения во времени вылупления и две мало различимые расы 17-летней цикады (Tibicen septemdecem L).
Во всех этих случаях едва намечающиеся различия сопровождаются значительной затрудненностью скрещивания между расходящимися формами вследствие разновременности созревания половых продуктов. Это — безусловно решающий момент, обеспечивающий фиксирование новых форм и их расхождение.
3. Физиологическая изоляция. Под физиологической изоляцией понимают затрудненность или малую эффективность скрещивания между представителями разных групп особей (несмотря на полную возможность их встречи).
В основе физиологической изоляции лежит как генотипическое и биохимическое, так и физиологическое и морфологическое расхождение признаков. В качестве последнего можно отметить расхождение в строении полового аппарта, которое делает копуляцию с исходной формой невозможной. Несомненно, такое расхождение иногда предшествует обособлению по другим морфофизио-логическим признакам (часто у насекомых). Однако оно все же представляет уже результат эволюции, связанной с взаимным приспособлением полов. Такое расхождение не может быть результатом единичной мутации (за исключением различий в общей величине тела, которые также иногда ведут к невозможности скрещивания), так как взаимность приспособления требует сложного согласования ряда отдельных изменений у обоих полов. То же самое касается видовых отметин и вторично-половых различий, которые должны быть также согласованы с изменениями в инстинктах и рефлексах, ведущих к спариванию. И такая форма физиологической изоляции вполне реальна (часто у насекомых). У высших животных она приобретает формы психической изоляции. У птиц она связывается главным образом со зрительными впечатлениями от «видовых отметин», частью со слуховыми восприятиями (пение птиц). У млекопитающих основную роль играет расхождение в пахучих железах, что связано с высоким развитием органа обоняния и с его ролью в половом возбуждении. И здесь условия взаимности и согласованности в изменениях видовых отметин (окрасок, пахучих желез) с изменениями в инстинктах и поведении несовместимы с представлением об единичных мутациях или даже о простейших их комбинациях. И эти изменения мы должны рассматривать как результат эволюционного процесса, не начинающего, а завершающего видообразование. Поэтому мы оставляем здесь эти классические формы физиологической или половой изоляции без рассмотрения,— хотя возникновение половой изоляции и падает иногда на расовые различия, как это доказано, например, для двух рас (А и В) Drosophila pseudoobscura (Lancefield, Boche, по Добжанскому, 1937),— а фиксируем все наше внимание на других формах изоляции, которые, быть может, имеют несколько большее значение если не в самом возникновении расовых различий, то по меньшей мере в их относительно раннем фиксировании в качестве жизненной, в данных условиях, комбинации наследственных изменений. В этом смысле имеют значение биохимическое и генетическое расхождения, которые могут быть обозначены как гаметная и генетическая формы изоляции.
Гаметная изоляция. Межвидовая гибридизация остается обычно безрезультатной и при отсутствии каких-либо механических препятствий и даже при искусственном осеменении животных или опылении растений. У растений пыльца на чуждом рыльце не прорастает, пыльцевая трубочка не проникает в ткань столбика, не доходит до зародышевого мешка или, даже при ее успешном прорастании, генеративная клетка все же гибнет, оставляя яйцеклетку неоплодотворенной. У животных сперматозоид не проникает в яйцо или гибнет в его протоплазме. Иногда яйцо начинает дробиться, но хромосомы сперматозоида не участвуют в дроблении, выбрасываются из ядра и дегенерируют.
Во всех этих случаях мы имеем явные выражения какой-то, очевидно биохимической, несовместимости сперматозоида с тканями чуждого организма и несовместимости ядра сперматозоида с плазмой яйцеклетки. В других случаях оплодотворение совершается и дробление начинается нормально, но зародыш гибнет. У животных это происходит, по-видимому, чаще всего непосредственно после гаструляции (во время которой протекают важнейшие детерминационные процессы). В этом случае биохимическое расхождение, очевидно, не столь велико, чтобы вызвать гибель ядерных субстанций в чуждой плазме яйца, но все же настолько значительно, что их взаимодействие в процессах обмена (и в- стоящих с ними в связи детерминационных процессах) невозможно. Иногда гибридные зародыши вначале развиваются нормально и гибнут на более поздних стадиях, либо дают начало очень слабой молоди. Смертность среди таких гибридов очень велика.
Все это представляет очевидный результат биохимического расхождения организмов, в котором нарушается нормальное взаимодействие ядерных веществ и плазмы, а следовательно — нарушается и нормальное течение процессов обмена, лежащих в основе индивидуального развития и всех вообще жизненных процессов организма. Поскольку это расхождение проявляется уже в большей или меньшей несовместимости половых клеток, можно говорить о гаметной изоляции.
Совершенно ясна постепенность этого расхождения, которое проявляется в полной мере лишь в межвидовой гибридизации (да и то не всегда — в некоторых случаях возможны и межвидовые гибриды и даже увеличение мощности межвидовых гибридов — явления гетерозиса). Тем большее значение имеет для нас то обстоятельство, что начальные фазы биохимического расхождения замечаются иногда также в расовых признаках.
По Дикте (Pictet, 1936), жизнеспособность гибридов между более отдаленными географическими расами дубового шелкопряда (Lasiocampa quercus) заметно снижена. То же самое наблюдалось еще резче при скрещивании местных рас другой бабочки (Nemeo-phila plantaginis), у которых иногда гибридизация совсем не удавалась вследствие отсутствия оплодотворения. Особенно интересно, что такое снижение жизнеспособности наблюдается иногда и при скрещивании различных мутаций дрозофилы. Некоторое снижение жизнеспособности видно на комбинации abnormes — lozenge у Drosophila funebris (Тимофеев-Ресовский, 19356). Более резко это выражено в комбинации curly-wing — moire eye (Мёллер).
Очень часто расхождение двух форм выражается в том, что у гибридов недоразвиваются половые железы и половые клетки в них не развиваются или дегенерируют. Так, например, даже при скрещивании различных рас Drosophila pseudoobscura (А и В), именно — при оплодотворении самки В самцом А, получаются стерильные самцы с редуцированными семенниками. Тщательный анализ таких, а также и обратных скрещиваний (когда получаются стерильные самцы с семенниками нормальной величины), проведенный Ф. Добжанским, показал, что эти дефекты являются результатом взаимодействия наследственного материала двух разных рас (при участии не менее 8 генов «стерильности»). При скрещивании различных рас непарного шелкопряда (Lymanthria dispar) получаются, как известно (Гольдшмидт, 1935), различные интерсексы, т. е. гибриды с ненормальным половым аппаратом. И здесь, как показал Р. Гольдшмидт, нарушения в развитии половых органов объясняются взаимодействием различного наследственного материала у гибридов «слабых» и «сильных» рас этого шелкопряда. В этих последних случаях, когда недоразвивается половая система, мы имеем переходы к следующей форме — генетической изоляции, для которой характерна стерильность развивающихся гибридов. Именно в этих явлениях мы имеем начальные фазы расхождения. И в этом опять подчеркивается значение наследственных изменений ядерных структур во всех процессах внутривидового дифференцирования. Несовместимость ядра и плазмы сказывается, обычно, лишь на межвидовой или даже межродовой гибридизации, и это заставляет нас еще раз вспомнить сказанное ранее об изменчивости и, следовательно, малой устойчивости ядерных структур и относительно большей устойчивости плазмы.
Генетическая изоляция. Еще Федерлей (Federley, 1913–1916) показал на целом ряде скрещиваний различных видов бабочек (виды Pygaera и др.), что стерильность гибридов обусловлена расстройством редукционных делений при образовании половых клеток. Во время мейозиса чуждые хромосомы, все или некоторые, не спариваются и не образуют нормальных бивалентов. Эти данные были затем подтверждены и дополнены другими исследованиями над межвидовыми гибридами как у животных, так и у растений. Неконъюгирующие хромосомы (униваленты) распределяются случайно, так что половые клетки получают ненормальное и притом различное число хромосом. Важно отметить, что уже само накопление генных различий ведет к уменьшению синаптиче-ского сродства хромосом (Мёллер). Керкис (1936) нашел, что у гибридов D. melanogaster X D. simulans гомологичные хромосомы конъюгируют (в слюнных железах) совершенно даже в тех участках, которые имеют одинаковое строение. То же найдено у гибридов Drosophila virilis X D. americana (Hughes, no Muller, 1940).
Очевидно, ослабление притяжения связано с различиями в самих генах. Однако дело не исчерпывается одними лишь ненормаль-ностями в конъюгации и распределении хромосом. Клетки с нарушенным «балансом» хромосом вследствие ненормального их распределения оказываются нежизнеспособными. Что здесь дело именно в нарушении баланса, видно из того, что если у межвидовых гибридов произойдет удвоение числа хромосом, так что каждая хромосома получает свою пару, то такие «аллотетраплоиды», имеющие полные наборы хромосом, оказываются вполне плодовитыми и бойкими при внутреннем размножении. Впрочем, иногда хромосомы образуют правильные пары, но все же проявляются ненормальности в процессах редукционного или эквационного деления, и половые клетки после этого дегенерируют (иногда они и вообще недоразвиваются и не доходят даже до редукционных делений, как мы отмечали при рассмотрении гаметной изоляции). Все это относится к межвидовым гибридам. Нас интересует здесь, однако, лишь начальная дифференциация форм внутри вида и, следовательно, для нас имеет особое значение вопрос о возникновении механизма генетической изоляции и о его возможной роли в этом процессе внутривидовой дифференциации.
Ф. Добжанский (1937), который обстоятельно разработал вопрос о возникновении стерильности, различает генную и хромосомную стерильность. Нам незачем входить в особые детали этого вопроса, однако отметим, что иногда стерильность определяется изменением одного лишь гена, который обнаруживает типичное менде-лирование при скрещивании. Несколько таких генов описано Бидлем у кукурузы (Beadl, 1930–1933). Описаны такие гены и для видов дрозофилы (Sturtewant, 1929 а, б; Gowen, 1931). Здесь отдельная мутация выражается в нарушениях конъюгации хромосом, в их неправильном расхождении или в других ненормально-стях при образовании половых продуктов. В высшей степени интересны данные Мёллера и Раффеля, показавшие, что при скрещивании двух вполне плодовитых мутаций дрозофилы curly-wing и moire eye получаются слабые и бесплодные гибриды. То же самое наблюдается при скрещивании scute-18 и scute-4 (Muller, 1940). Здесь получаются, следовательно, стерильные комбинации о т-дельных мутаций. Чаще всего, по-видимому, стерильность определяется не отдельной мутацией, а несовместимостью целых генотипов, которые дают негармоничную комбинацию, ведущую, по меньшей мере, к бесплодию. В других случаях стерильность определяется несовместимостью хромосом вследствие наличия крупных структурных различий — именно транслокаций или более сложных инверсий. Расы А ж В D. pseudoobscura различаются четырьмя инверсиями. При скрещивании получаются стерильные самцы. Кожевников (1936) получил расу дрозофилы {D. melanogaster), отличающуюся двумя транслокациями (D. artificialis). При чистом разведении она обладает лишь сниженной плодовитостью (жизнеспособна lU часть всех гамет). Скрещивание с исходной нормальной дрозофилой остается безрезультатным, так как оно не может привести к образованию зиготы со «сбалансированной» системой хромосом.
Генетики считают, что в основе стерильности гибридов лежит либо изменение некоторых генов, т. е. отдельные мутации «стерильности», которые обнаруживают свое влияние в чуждом генотипе, либо негармоническая комбинация генов или хромосом (т. е. нарушение их «баланса»).
Хотя физиологическая изоляция является, очевидно, дальнейшим и притом обычно побочным результатом эволюции, интересно отметить, что все же она и сама по себе может быть результатом непосредственного действия естественного отбора. Два близких вида дрозофилы D. pseudoobscura и D. miranda могут быть скрещены, но дают стерильных гибридов. Между ними имеется известная физиологическая изоляция — в смешанных популяциях (в эксперименте) преобладают, особенно первое время, внутривидовые спаривания. Эта изоляция различно выражена для различных рас (раса В. у D. pseudoobscura) и линий (D. miranda) l, которые обитают в одном и том же ареале, и значительно ниже для тех рас и линий, которые происходят из отдаленных местностей (Добжанский, 1937). Очевидно, шел естественный отбор на физиологическую изоляцию только тех популяций различных видов, которые, живя в одной местности, фактически могли встречаться. Этот отбор основан на элиминации потомства особей, которые скрещивались с особями другого вида (так как это скрещивание вело к образованию бесплодных гибридов).
Накопление генных мутаций и хромосомных перестроек ведет к уменьшению сродства, выражающемуся в уменьшении притяжения между гомологичными хромосомами. Оно ведет к нарушению редукционных делений и к другим проявлениям несовместимости наследственного аппарата расходящихся форм. Нарушение «баланса» генов и хромосом является таким же проявлением их несовместимости в процессах обмена веществ и в индивидуальном развитии организма. Оно ведет к «летальности» или «стерильности» несовместимых комбинаций. Несовместимость ядра и плазмы является дальнейшим и еще более глубоким проявлением биохимического расхождения различных форм в процессе эволюции. Все эти процессы способствуют все более полному их обособлению.
Колебания численности популяций (периодическое ограничение панмиксии). Мы уже говорили вначале о роли чередования различных форм размножения, связанных с периодическим прекращением панмиксии. У высших животных такое чередование поколений не играет никакой роли (за немногими исключениями), но зато у многих животных наблюдается временное ограничение панмиксии, связанное с более или менее резким сокращением численности. Мы знаем, что панмиксия осуществляется более свободно лишь в больших популяциях подвижных животных. В малых популяциях преобладает родственное скрещивание, ведущее к гомозиготизации и закреплению различных случайных комбинаций наследственных изменений. В первом случае (в больших популяциях) свободная панмиксия позволяет осуществить много новых комбинаций, но не дает возможности их закрепления. Во втором случае (в малых популяциях) родственное скрещивание ограничивает возможность комбинирования, связано даже с утерей многих уже осуществленных комбинаций, но, с другой стороны, может привести к фиксированию и размножению некоторых благоприятных сочетаний. В частичной пространственной изоляции отдельных популяций широко распространенного вида происходит известное совмещение преимуществ большой популяции, создающей новые комбинации благодаря возможности миграций, и выгод, связанных с дифференциацией на малые популяции, что способствует закреплению удачных комбинаций.
При периодических колебаниях численности также происходит подобное совмещение обоих условий видообразования путем чередования процессов комбинирования и процессов фиксирования и размножения.
Колебания численности могут носить несезонный характер, и именно эти колебания имеют большое народнохозяйственное значение, так как их нелегко предусмотреть. Факторы, способствующие массовому размножению, и факторы, определяющие наступление следующего за ним периода депрессии, могут быть весьма различными. Это могут быть случайные изменения в физических факторах (температура, влажность), прямо или косвенно благоприятствующих размножению или, наоборот, ограничивающих это размножение, изменения в условиях питания (неурожаи кормов) или изменения в биологических соотношениях (истребление хищниками, истощение паразитами, эпизоотии). Иногда выясняется и вполне закономерная зависимость в колебаниях численности различных организмов, объединенных пищевыми связями. Известны сопряженные изменения численности зайца и рыси в Канаде (Hewitt, 1921) и многие другие связи, прослеженные на протяжении многих лет главным образом по масштабам заготовок шкурок пушных зверей, а также по биологическому учету численности (см. сводки С. А. Северцова, 1941; А. Н. Формозова 1934; Elton, 1924, 1927). Несезонные колебания численности насекомых, а отчасти грызунов и даже крупных млекопитающих могут определяться и метеорологическими условиями — случайно особо благоприятная весна может привести к интенсивному размножению насекомого, дождливое лето может значительно снизить его численность, а глубокий снежный покров и, в особенности, его обледенение могут привести к длительной голодовке и массовой гибели даже крупных травоядных млекопитающих. Это все — наиболее простые случаи. Чаще всего, однако, у мелких животных (насекомых), при сложности связей и малой специализации питания, массовое размножение или почти полное исчезновение зависит от сложного сочетания многих благоприятных или неблагоприятных факторов.
Для процессов эволюции имеют, пожалуй, еще большее значение почти всеобщие для умеренных и холодных стран явления сезонных колебаний численности. Во время зимовок ежегодно гибнет большое число птиц и млекопитающих и огромное количество насекомых. Отношение численности популяций перед зимовкой и после нее достигает 1000 :1 или более. Разница в величинах осенней и весенней популяций бывает очень значительной и у птиц, и у мелких млекопитающих.
Значение всех этих колебаний для эволюционного процесса станет для нас яснее, если мы учтем следующее. В период, благоприятный для размножения животного, острота естественного отбора значительно падает, элиминация вредных мутаций становится менее интенсивной и поэтому идет усиленное их накопление, распространение и комбинирование. Индивидуальная изменчивость достигает известного максимума.
Однако с увеличением плотности населения увеличивается активная конкуренция особей за жизненные средства, а также начинается усиление деятельности хищников и паразитов, истребляющих данную форму. Борьба за существование обостряется и в большой популяции идет интенсивный и эффективный отбор наиболее активных, вооруженных и защищенных особей, т. е. наиболее благоприятных наследственных комбинаций. Вслед за этим дальнейшее усиление истребляемости ведет к более или менее резкому уменьшению численности. Интенсивные формы элиминации могут утратить избирательный характер. Это касается в особенности действия стихийных для данного организма сил, как эпизоотии, некоторые паразитарные заболевания, зимний холод или особо мощные (по сравнению с жертвой) хищники. В таком случае переживание может приобрести до известной степени случайный характер. Однако в масштабах целого вида и значительных промежутков времени вряд ли случайные явления играют большую роль. Численность популяций достигает известного минимума и вместе с тем увеличивается ее генетическая однородность. Некоторые популяции, пережившие оба периода борьбы и в большинстве особей оказавшиеся благоприятными сочетаниями генов, сохраняются, другие — теряются. Однако случайности утери и сохранения различны в различных местных популяциях и поэтому повсеместная утрата наиболее благоприятных сочетаний совершенно невероятна. Во всяком случае, в периоды депрессии происходит закрепление некоторых комбинаций, а в периоды следующей затем экспансии эти комбинации размножаются и могут достигнуть большой численности.
Таким образом, и в периодических колебаниях численности животных имеются условия, с одной стороны, облегчающие установление, размножение и отбор новых комбинаций (в процессе панмиксии в большой популяции) и, с другой стороны, облегчающие фиксирование некоторых благоприятных комбинаций при резком сокращении численности и их размножение при новом увеличении численности.
Что это не отвлеченные предположения, видно по целому ряду фактов.
Н. Ford и Е. Ford (1930) наблюдали в течение большого ряда лет ограниченную и изолированную популяцию бабочки Ме-litaea aurinia. При резком уменьшении ее численности популяция имела весьма однородный характер. После этого быстрое увеличение численности привело к значительному усилению изменчивости. Потом численность популяции стабилизировалась и изменчивость опять снижалась. Популяция стала вновь однородной. Новая норма оказалась, однако, заметно отличной от той, которая была в предыдущем стабильном периоде существования популяции.
Целый ряд исследований показал ясную зависимость между сезонными изменениями численности и самой изменчивостью. Томпсон, Белл и Пирсон (Thompson, Bell, Pearson, 1911) сравнивали изменчивость осенней и весенней популяций обыкновенной осы. Зимнее отмирание популяции привело к значительному сокращению ее изменчивости. Лукин (19396) исследовал популяцию клопов-«солдатиков» (Pyrrhocoris apterus) и установил у них существование как локальной, так и сезонной изменчивости размеров тела. По Калабухову (1941), исследовавшему на Сев. Кавказе популяцию домашней мыши, в годы низкой численности (1927–1931) она состояла в основном из светлобрюхих особей, типа курганчиковой мыши, а в 1933 г. при значительном увеличении популяции наблюдались самые различные окраски, в том числе 25–30% очень светлых.
Р. Л. Берг установила, что во время летнего увеличения численности популяций дрозофилы их насыщенность мутациями значительно возрастает..Уже осеннее сокращение популяции ведет к значительному снижению числа встречающихся мутаций. То же самое можно установить для мигрировавших популяций (отбор во времени миграции). Деятельность отбора в сокращении изменчивости видна и по другим наблюдениям того же автора. Среди мух, только что вылупившихся из куколок, аберраций оказывается больше, чем среди взрослых мух той же популяции. Все эти факты показывают, что количество видимых изменений тем меньше, чем интенсивнее отбор в данных условиях (Р. Берг, 1943а, б).
Зимнее сжатие популяции ведет затем к дальнейшему сокращению изменчивости. Здесь элиминация теряет свой избирательный характер и при стихийности элиминирующих факторов освобождает значительное место также и случайным явлениям. Наличие таких явлений — в результате как локального, так и сезонного ограничения популяций — было установлено Н. Дубининым на той же дрозофиле (Дубинин, 1931; Дубинин и Ромашов, 1932, Дубинин и др., 4934).
9. РЕЗЕРВ НАСЛЕДСТВЕННОЙ ИЗМЕНЧИВОСТИ ВИДА И ЕГО МОБИЛИЗАЦИЯ
Взаимодействие всех разобранных процессов — постоянного мутирования, элиминации вредных и распространения безвредных или условно и частично вредных мутаций через процесс размножения, их разнообразного комбинирования в результате свободного скрещивания (панмиксии), под непрерывным контролем естественного отбора, поощряющего распространение наиболее благоприятных комбинаций, и закрепления некоторых комбинаций в условиях локального или временного ограничения панмиксии — ведет к накоплению внутри любой популяции известного запаса индивидуальной изменчивости. Такой внутривидовой запас изменчивости определяет эволюционную пластичность данной популяции при изменении соотношений между организмом и факторами внешней среды. Как видно, это накопление резерва изменчивости зависит от многих факторов: скорости мутирования, остроты элиминации и интенсивности естественного отбора, а также от свободы комбинирования, зависящей, в свою очередь, от величины популяции, степени ее изолированности и т. д. Поэтому в различных условиях борьбы за существование и эволюционная пластичность разных популяций и разных видов организмов бывает весьма различной.
А. УСЛОВИЯ НАКОПЛЕНИЯ РЕЗЕРВА ИЗМЕНЧИВОСТИ (В ПОПУЛЯЦИЯХ)
Значение мутабилъности. Лишь при непрерывном появлении все новых мутаций они могут накапливаться в популяции. Единичные мутации обычно теряются, даже и в том случае, если они благоприятны для данного вида организмов в данных условиях. С другой стороны, повторно возникающие мутации могут при достаточно высоком мутационном «давлении» накапливаться в популяции до известной степени и в том случае, если они неблагоприятны. Они могут преодолеть «давление» отбора вплоть до достижения уровня равновесия, определяемого интенсивнос-тями обоих противоположных процессов.
Существуют как высоко мутабильные популяции и линии, так и мало мутабильные. По Р. Л. Берг, мутабильность обыкновенной дрозофилы (Drosophila melanogaster) из больших популяций Умани и Никитского сада в Крыму очень высока; мутабильность той же дрозофилы из малых популяций Дилижана (Армения) и Каширы (Московская область) очень низка. Низка также мутабильность американской линии «Флорида». Однако и в популяциях с низкой мутабильностью встречаются отдельные очень высоко мутабильные линии (Demerec, Шапиро, Берг).
Встречаются и мутации, увеличивающие темп мутирования. Мутабильность оказывается таким же наследственным признаком, меняющимся в процессе мутирования, как и любые другие признаки организма. Генетики говорят и о «генах» мутабилъности, так же как о «летальных генах» или о «генах» стерильности. Само собой разумеется, что это лишь упрощенный способ выражения той мысли, что некоторые наследственные изменения одного «гена» вносят столь значительное расстройство в координи-рованность морфофизиологических (включая биохимические) процессов, что делают организм нежизнеспособным, делают невозможным его развитие (летали), прекращают развитие половых желез или половых клеток (гены «стерильности»). Иногда же эти изменения лишь немного нарушают «баланс» наследственного аппарата так, что он оказывается менее устойчивым и в нем легче возникают новые мутации (гены «мутабилъности»).
Значение темпа мутирования для скорости распространения мутаций в популяции ясно само собой. Для нас важно, что при прочих равных условиях, т. е. прежде всего при равной интенсивности элиминации, повышенная частота возникновения мутаций («мутационное давление») ведет к увеличению концентрации мутаций и к установлению равновесия на более высоком ее уровне. Таким образом, в высоко мутабильных популяциях и насыщенность их мутациями должна достигать более высокого уровня. Мы уже видели, что она для неблагоприятных мутаций достигает равновесия на уровне, пропорциональном частоте мутирования (и) и обратно пропорциональном коэффициенту селекции (s) в пользу нормы, согласно формуле для нормы q — 1 — u/s, а следовательно — для мутации: (1 — q — u/s).
Изменение мутабильности является, очевидно, лишь косвенным результатом эволюции. При обострении борьбы за существование и увеличении интенсивности избирательной элиминации, — особенно в относительно постоянных и однородных условиях внешней среды, —- всех более значительных уклонений от приспособленной нормы (стабилизирующая форма отбора) происходит сокращение изменчивости, т. е. «иммобилизация» вида. Косвенным результатом элиминации большого числа мутаций является преобладающая элиминация мутабильных линий, как дающих максимум мутаций, и, следовательно, преобладающее переживание более однородного («нормального») потомства менее мутабильных линий.
Обратный процесс повышения мутабильности может быть результатом сохранения вновь возникающих линий с более высокой мутабильностью (мутации «мутабильности») в силу ослабления интенсивности элиминации, при уменьшении остроты борьбы за существование, особенно в условиях разнообразной и меняющейся внешней среды. Возникновение таких линий в результате отдельных мутаций наблюдалось неоднократно (Н. Шапиро, 1938). Простое их сохранение при благоприятных условиях ведет к увеличению мутабильности популяций через распространение этих мутаций до нового уровня равновесия (определяемого хотя и косвенно, но в конце концов также через соотношение u/s).
В некоторых случаях повышение мутабильности может быть результатом и другого процесса. В межгрупповой конкуренции при меняющихся условиях среды побеждают те популяции, которые быстрее приспосабливаются к новым условиям существования. При прочих равных условиях побеждать будут более мута-бильные популяции и более мутабильные линии. Таким образом, мутабильность может меняться в результате межгруппового соревнования в темпах эволюции (Р. Берг). Не отрицая значения межгруппового соревнования в определении мутабильности, следует отметить, что указанный ранее фактор — интенсивность элиминации мутаций, а следовательно и мутабильных линий — должен быть значительно более эффективным. К тому же он независим от темпов эволюции и скорости изменения внешней среды. Интенсивность элиминации определяется лишь условиями борьбы за существование, и это вполне объясняет наличие существующих в настоящее время различий. В однородных условиях существования малых популяций преобладает иммобилизирующая роль отбора (острая элиминация уклонений) и, следовательно, преимущественное распространение менее мутабильных линий. В разнородных и благоприятных условиях существования больших популяций, с возможностью широких миграций, элиминация уклонений не столь остра и вполне возможно общее повышение уровня изменчивости, а вместе с тем и сохранение более мутабильных линий.
Б. СОСТАВ МОБИЛИЗАЦИОННОГО РЕЗЕРВА
Вредные мутации и комбинации постоянно элиминируются, а полезные быстро завоевывают всю популяцию. Поэтому мобилизационный резерв видов и популяций составляется в основном из «нейтральных» мутаций и «сбалансированных», т. е. из безразличных и, быть может, частично благоприятных их комбинаций. Вопрос о самом существовании вполне нейтральных мутаций подвергается справедливому сомнению. Что же касается неблагоприятных мутаций, то они, казалось бы, и не могут накапливаться. Не нужно, однако, подходить слишком формально к пониманию «нейтральности» мутаций или их комбинаций. К нейтральным мутациям следует причислить все мутации, которые не получают заметного выражения вследствие наличия совершенного регуля-торного аппарата, сглаживающего нарушенные соотношения.
К категории нейтральных мутаций можно отнести прежде всего вообще все малые мутации, обладающие столь незначительным выражением, что они не могут обусловить заметного снижения жизнеспособности их обладателей. Предполагается, что малые мутации возникают чаще. Однако, если это даже и неверно, то насыщенность природных популяций малыми мутациями, во всяком случае, очень высока. Многообразие выражения многих мутаций, которое не только в разных расах, популяциях и линиях (Тимофеев-Ресовский, Добжанский, Дубинин), но даже у отдельных особей оказывается различным; неполнота проявления многих мутаций даже в строго контролируемых идентичных условиях культуры,— все это указывает на существование многих небольших, трудно анализируемых, но именно генотипических различий между популяциями, между разными линиями и даже отдельными особями наших обычных культур. Экспериментально это может быть продемонстрировано внесением определенной мутации (например, beaded) в различные линии или успег.шыми результатами искусственного отбора на максимальное или минимальное выражение известного признака (Камшилов, 1939; Шифрин, 1941). В особенности изменчиво выражение многих мутаций в гетерозиготном состоянии. Быстрота, с какой меняется это выражение в лабораторных линиях, показывает высокий уровень насыщения малыми мутациями даже чистых культур, во всяком случае — численно очень ограниченных, экспериментальных популяций. Значение малых мутаций в эволюции также бесспорно.
Во-вторых, к категории нейтральных мутаций можно отнести и более значительные мутации, если они в гетерозиготном состоянии заметно не нарушают общей жизнеспособности организма. Это касается главным образом рецессивных и полудоминантных мутаций. Так как первоначальное накопление мутаций касается всегда гетерозиготы, то все рецессивные (даже вредные) мутации распространяются совершенно свободно. Однако новые мутации, как правило, полудоминантны (Фишер, 1931) и, следовательно, имеют некоторое выражение и в гетерозиготе. Если это выражение не слишком неблагоприятно, по крайней мере в некоторых генотипах, то и такие мутации могут накапливаться. В процессе их накопления и комбинирования они могут потерять свои вредные выражения (через отбор малых мутаций — модификаторов). Природные популяции дрозофилы действительно насыщены и вредными мутациями не только рецессивными, но и полудоминантными, выражения которых могут быть подавлены модификаторами, имеющими частично эпистатическое влияние (Берг, Гершензон).
Можно считать доказанным, что в состав резерва наследственной изменчивости входят и более крупные мутации, вредные выражения которых частью погашены в процессе подбора модификаторов. Это — большая категория нейтрализованных или обезвреженных мутаций, которые по мере их нейтрализации могут достигнуть все более высокого уровня насыщения популяции. В этом случае, как и в предыдущем, популяция может быть по внешности весьма однородной. Индивидуальные различия очень невелики, так как выражения отдельных мутаций в значительной мере погашены.
В-третьих, мы можем условно причислить к той же категории нейтральных мутаций, входящих в состав мобилизационного запаса, и частично вредные (частично полезные) мутации, которые, будучи в некоторых отношениях неблагоприятными, оказываются в других отношениях (на других стадиях или в других своих выражениях) благоприятными. Такие мутации для организма в целом оказываются действительно нейтральными. В процессе эволюции вредные выражения таких мутаций будут погашаться (подбором модификаторов) и накопление их может идти возрастающими темпами по мере того, как они становятся все более благоприятными.

Рис. 18. Гетероморфизм окраски у божьей коровки Adalia bipunctata. Изображены только гомозиготные формы. По Я. Я. Лусу, 1932
В-четвертых, к тем же нейтральным мутациям мы отнесем и мутации условно вредные (условно полезные). Если популяция живет в разнородной среде, то локальные условия существования предъявляют различные требования к организму. Организм, вполне приспособленный к одним локальным условиям, оказывается менее приспособленным к другим локальным условиям. Мутация, неблагоприятная в одних условиях (например, мелани-стическая форма на светлом фоне), может оказаться благоприятной в других (та же меланистическая форма на темном фоне).
Гетероморфизм. При наличии известного, более или менее постоянного, разнообразия условий, характерного для данного ареала распространения, возможно установление локального, более или менее постоянного соотношения разных форм. Одни и те же наследственные формы входят в состав различных популяций данного вида в различном процентном соотношении. Таково, например, географическое распределение разных типов окраски у божьих коровок (Harmonia oxyridis, по Добжанскому, 1937, и другие виды — рис. 10, 18). Подобным же образом весьма постоянное соотношение имеется между разными типами полосатых и гладких (т. е. однотонных) раковин садовых улиток (Cepea hortensis и С. nemoralis). Это соотношение мало меняется даже в течение геологических периодов (Diver, 1929, по Холдену, 1935). Такую форму генетического полиморфизма можно обозначить термином равновесный гетероморфизм. Еще ярче выражен такой гетероморфизм у обыкновенного богомола (Mantis religiosa). Он характеризуется одновременным существованием различных ясно выраженных защитных окрасок (рецессивной — зеленой, и доминантной — буро-желтой) у отдельных особей в известном числовом соотношении (для Карадага в Крыму 1:2,5), что связано с их распределением на соответствующей растительности (Беляев, 1946). Зеленые держатся на зеленых растениях (в Карадаге на каперцах), а буро-желтые — на высохших. Подобный же равновесный гетероморфизм ограничивается иногда одним полом, как в случае мимикрии у самок Papilio dardanus и P. polytes, генетический механизм которого разобран Фишером (1930).
Внутривидовой гетероморфизм имеет приспособительное значение. Каждая форма в целом все полнее приспособляется к известным частным условиям существования. Таким образом, возможна дальнейшая дифференциация вида на локальные формы (экотипы). Для разбираемых здесь вопросов важно, что в случае изменения внешней среды (ее климатических или биотических факторов) условия, бывшие прежде случайными или локальными (засуха, холод, недостаток света, влажность, размножение хищников или организмов, служащих пищевым материалом), могут стать более частыми, постоянными или всеобщими. В ответ на такое изменение в данной популяции произойдет в основном лишь сдвиг в сторону преобладания одной из ранее существовавших форм, которая теперь получила постоянное преимущество. Малочисленная второстепенная форма может быстро стать преобладающей. В таких случаях можно говорить об известной степени «пре-адаптации» данного вида к новым условиям в некотором числе особей. Такая преадаптация существует, однако, только благодаря тому, что эти новые условия встречались уже и раньше в жизни вида, но не как постоянные, а как случайные, периодические или локальные особенности существования.
Несомненно, что известный гетероморфизм, постоянно наблюдаемый в популяциях вида, живущего в разнообразных условиях внешней среды, значительно повышает его эволюционную пластичность. Возникая на базе индифферентного мобилизационного резерва мутаций, накапливающихся в популяции и комбинируемых с тем большей скоростью, чем многочисленнее популяция и чем более разнообразны условия внешней среды в пределах ареала ее распространения, этот гетероморфизм еще до распада вида на экотипы и географические расы начинает приобретать приспособительное значение в известных условиях существования. В этом случае довольно быстрое изменение в факторах внешней среды может привести к очень быстрой мобилизации внутренних ресурсов и быстрому преобразованию вида в процессе эволюции. Чем больше запас индивидуальной изменчивости и генетический полиморфизм популяций, тем выше эволюционная пластичность вида.
Обычный резерв индивидуальной изменчивости имеет в моно-морфном виде в значительной мере скрытый характер — по внешности особи могут быть весьма однородными («дикий» тип дрозофилы) даже и в больших популяциях.
В. УСЛОВИЯ ВСКРЫТИЯ И МОБИЛИЗАЦИИ РЕЗЕРВОВ
Мобильность видов означает их потенциальную способность к эволюции. Эта потенциальная способность может быть реально использована лишь в известных условиях. Естественный отбор имеет полем своего приложения всегда лишь реализованные фенотипы, а не потенциально возможные. Поэтому непосредственным материалом для движущей формы отбора и для прогрессивной эволюции могут быть лишь мутации и их комбинации, имеющие какое-либо физиологическое или морфологическое выражение, а не скрытые формы мутаций. Лишь явная наследственная гетерогенность популяций, т. е. наличие индивидуальных уклонений или зарождающихся форм групповой изменяемости (гетероморфизм), может быть непосредственно использована в эволюции организмов. Все отрегулированные или нейтрализованные формы изменчивости, входящие в состав «мобилизационного резерва», должны быть сначала выявлены, мобилизованы и лишь после этого могут стать предметом прямого отбора (в его ведущей форме). Таким образом, первым ответом вида на изменение внешней среды, или точнее — на изменение соотношений между организмом и внешней средой, является более или менее быстрое вскрытие его резервов изменчивости.
а. Вскрытие резервов изменчивости. Вскрытие резервов осуществляется прежде всего в процессе гомозиготизации мутаций и их комбинаций, которые в новых условиях существования оказываются благоприятными. В более широком масштабе это возможно при разбивке вида на малые изолированные популяции, разбивке, ведущей к родственным скрещиваниям. Гомозиготиза-ция может принять всеобщий характер при переходе к апогамии, что наблюдается нередко у растений. В этом случае очень ясно проявляется гетерогенность популяций (одуванчик, ястребинка, крупка и др.).
Второй путь лежит через индивидуальное приспособление и снятие в связи с этим некоторых регуляций. Изменение внешних факторов может и прямо коснуться регуляторного аппарата, всегда отличающегося известной лабильностью, и несколько нарушить полноту регуляции. С другой стороны, еще важнее, что в процессе индивидуального приспособления меняются функциональные соотношения частей, т. е. теряют свое значение некоторые прежние соотношения и устанавливаются новые. Прежняя «регуляция» теряет свою координированность и снимается в том же порядке индивидуального взаимоприспособления частей, в каком она в свое время возникла. Это будет способствовать выявлению таких наследственных изменений, которые ранее сглаживались путем различных функциональных компенсаций. В новых условиях эти компенсации просто отпадают.
Изменение внешних факторов может и непосредственно повлиять на выражение существующих мутаций. Это выражение было погашено лишь в прежних условиях развития организма и может так или иначе проявиться в новых условиях (мы знаем, что выражение мутации зависит как от генотипа организма, так и от внутренних факторов его развития и, наконец, и от факторов внешней среды, см. 1–4). При воздействии внешних факторов (повышенной температуры, по Child, Blanc and Plough, 1940) на гетерозиготу нередко выявляются признаки даже вполне рецессивной мутации. Нередко отмечалась повышенная изменчивость популяций, живущих в неблагоприятных условиях (при крайне высоких или низких температурах и т. п.).
Наконец, важнее всего то обстоятельство, что каждая новая мутация прямо или косвенно способствует выявлению имеющихся уже скрытых, а также и любых новых мутаций.
Некоторые мутации настолько изменчивы в своих выражениях, что даже употреблялись как индикаторы для анализа генетического состава линий и популяций (Тимофеев-Ресовский, Добжанский, Дубинин). У дрозофилы это касается многих мутаций, характеризующихся изменением числа щетинок или жилкования крыла, а также мутаций, выражающихся в уменьшении числа фасеток глаза. В различных генотипах они получают различное выражение, и если в нормальном генотипе они утратили свое выражение, то в любом ином они могут его восстановить. Такое восстановление наблюдалось при скрещивании лабораторных линий мутантов, утративших свое выражение (eyelles, stubbloid и др.). Реактивная способность тканей может быть вполне нормальной в довольно широких пределах между нижним и верхним порогами их раздражимости. Однако каждая мутация, сдвигающая норму реакции или меняющая концентрацию формативного раздражителя, приближает тот или иной порог нормальной реактивности. Поэтому другие мутации, которые в норме укладывались бы еще в рамки нормальной реактивности, могут у такого мутанта выйти за эти пределы. Приближение уровня фактического осуществления реакции к одному из порогов нормальной реактивности означает уменьшение «защищенности» морфогенетической реакции (уменьшение «морфогенетического запаса», по М. Камшилову). Малейший новый сдвиг в том же направлении нарушает ее нормальное течение и проявляется в виде более или менее заметного изменения строения и функций организма.
Поэтому каждая мутация, сдвигающая норму реакции в известном направлении, способствует выявлению всех мутаций, влияющих в том же направлении. Так как любая мутация обладает плейотропным выражением (хотя и добавочные выражения могут быть подавлены), то это касается не одной реакции, а в большей или меньшей мере целого ряда формообразовательных процессов. Можно смело сказать, что каждая мутация, вызывая сдвиг во времени и скорости возникновения, а следовательно — в интенсивности действия формообразовательного фактора, или, наоборот, вызывая сдвиг в реактивной способности тканей (практически это одно и то же), тем самым подготовляет почву и облегчает выявление любой старой или новой мутации, вызывающей сдвиг в том же направлении. По всей вероятности, это наиболее существенный фактор, способствующий быстрому вскрытию резервов при изменении условий существования, при котором некоторые мутации оказываются благоприятными и размножаются при содействии естественного отбора (без погашения их положительных выражений). Чем большее число мутаций при этом используется, тем большее их число приобретает явный характер.
Вскрытие резервов мутационной изменчивости всегда сопровождается значительной лабилизацией формообразования и повышением общей реактивности организма.
Процесс вскрытия уже существующих резервов имеет гораздо большее значение, чем самая интенсивная мутабильность, так как мутации, ранее накопленные в скрытом виде, проявляются сразу в значительной концентрации, обеспечивая тем самым наиболее высокую эффективность естественного отбора.
Вскрытие резервов идет, однако, с наибольшей скоростью в тех случаях, когда начинается вообще довольно свободное накопление разнообразных мутаций. Это происходит при смягчении остроты избирательной элиминации (отбора), если вид переходит в благоприятные условия существования. В этом случае увеличение изменчивости сопровождается значительным возрастанием общей численности, расширением ареала распространения и может привести к быстрой прогрессивной эволюции (ароморфоз) или дальнейшей дифференциации (алломорфоз).
С другой стороны, и при обострении общей элиминации, когда она теряет свой избирательный характер (следовательно, тоже падает острота отбора), происходит также вскрытие резервов и значительное увеличение изменчивости. Оно, однако, сопровождается в этом случае уменьшением общей численности, устранением косвенных форм соревнования и преобладанием дегенеративных изменений (катаморфоз, Шмальгаузен, 1939а).
б. Мобилизация видов. Во всех случаях вскрытие резервов означает реализацию скрытых потенций, возможность использования латентных свойств вида в процессе его дальнейшей эволюции. Эта возможность тем выше, чем больше объем резерва, т. е. чем разнообразнее эти латентные свойства. Вид приобретает известную мобильность, которая может в благоприятных условиях и при эффективном естественном отборе привести к быстрой его перестройке. Повышение изменчивости связано с увеличением эффективности естественного отбора (пропорционально о2, согласно Фишеру) и, следовательно, с возможностью быстрой эволюции.
Решающим условием для вскрытия резервов и увеличения мобильности вида в известном направлении является изменение положения организма во внешней среде. Чаще всего это связано с изменением направления естественного отбора, при котором некоторые мутации и их комбинации получают известные преимущества перед нормой (ведущая форма отбора). Происходит гомози-готизация благоприятствуемых мутаций и дальнейшее вскрытие резервов, главным образом в той их части, которая лежит в направлении действия естественного отбора. Это может произвести впечатление направленного мутирования, тем более, что к такому частичному вскрытию резервов присоединяется и выявление новых мутаций в том же направлении (так как при приближении порогового уровня данных реакций они оказываются именно в этой области «незащищенными» каким-либо морфогенетическим «запасом»). Кроме того, мы знаем, что нередко возникновение одной мутации облегчает возникновение других аллеломорфов того же ряда, вплоть до достижения наиболее устойчивого (т. е. более вероятного) состояния, которое бывает как раз наиболее удаленным от прежней нормы (например, мутация белоглазия, white, у дрозофилы) . Это — обычная форма частичного вскрытия резервов и направленной мобилизации видов. Она вызывается таким же направленным изменением соотношений между организмом и внешней средой. Преобразование организма определяется ведущей формой естественного отбора и означает замену в процессе эволюции одних частных приспособлений — другими (алломорфоз, Шмальгаузен, 1939а).
Однако еще интереснее возможность общего вскрытия всех резервов и широкой мобилизации вида в любых направлениях. Это возможно в тех случаях, когда при изменении факторов среды сам отбор теряет свою строгую направленность или его влияние значительно ослабляется. Если организм теряет свою приспособленность, переходя в иную и притом разнообразную среду, в которой он встречает вместе с тем благоприятные условия для существования и размножения, то это может быть связано как раз с резким ослаблением интенсивности естественного отбора. Элиминация приобретает общий, т. е. главным образом случайный характер; в остальном она сводится лишь к гибели нежизнеспособных, т. е. негармонично построенных уклонений. Так следует представлять себе завоевание организмом новых и достаточно широких пространств в природе.
Так шло завоевание суши членистоногими и по этому же пути пошли затем первые наземные позвоночные (стегоцефалы и котилозавры). То же самое касается птерозавров и затем птиц, завоевывавших воздушную среду, водных рептилий и млекопитающих, вернувшихся в воду и нашедших здесь богатейший планктон и рыбное население океанских просторов, и многих, многих других случаев, различающихся лишь по масштабам. Образование нового водоема или острова означает освобождение нового места, которое может быть использовано с наибольшей эффективностью первыми вселенцами, если в этих местах условия делают их жизнь вообще возможной. Если условия очень благоприятны, то это ведет к бурному размножению, связанному с быстрой эволюцией, как это видно, например, по фауне Байкала с его бокоплавами, планария-ми, бычками и т. п. Однако речь идет, конечно, не только о завоевании территорий. То же самое касается и новых мест в «экономии» природы (что не всегда отделимо от понятия территории — в случае, например, воздушной или водной среды в целом), т. е. новых экологических ниш, которые не были ранее использованы или были использованы менее совершенными организмами, не могущими конкурировать с новой формой.
При ослаблении интенсивности естественного отбора, в благоприятных условиях существования, элиминация некоторых мутаций прекращается (или ослабевает) и это способствует выявлению множества «нейтральных» мутаций и их комбинаций, переходу их в гомозиготное состояние, появлению новых мутаций и более му-табильных линий. Это вскрытие резервов идет, однако, в этом случае не в одном направлении, а имеет всеобщий характер. Возрастает изменчивость популяциии во всевозможных направлениях.
По вопросу об изменчивости популяций в оптимальных условиях у экологов имеются большие разногласия. Это, несомненно, связано с неопределенностью самого понятия оптимальных условий и различием его критериев. Можно считать, что вид находится в оптимальных условиях: 1) в середине своего ареала, в котором распространение характеризуется максимальной сплошностью (биогеографический оптимум); 2) там, где население взрослых особей достигает максимальной плотности (экологический оптимум) ; 3) там, где смертность взрослых оказывается наименьшей и средняя длительность жизни наибольшей (физиологический оптимум) ; 4) там, где он наиболее обеспечен жизненными средствами и активная конкуренция сведена к минимуму, плодовитость максимальна, а элиминация имеет наиболее случайный характер. В данных оптимальных условиях все особи достаточно хорошо приспособлены, так как здесь среда не предъявляет каких-либо особых требований, кроме общей «жизнеспособности». Это последнее понимание оптимальных условий можно обозначить как биологический оптимум. В наших построениях мы оперируем с этим пониманием оптимума.
Если вид достигает в какой-либо области максимальной плотности населения (при минимальной смертности взрослых в результате малой прямой элиминации), то это означает обычно максимальную конкуренцию за жизненные средства, смертность молоди, периодическую голодовку, снижение плодовитости. Элиминация хотя и невелика, но имеет очень жесткий избирательный характер — элиминируются все уклонения от приспособленной нормы. Изменчивость в области такого оптимума может быть только минимальной (что нередко и устанавливалось).
Если мы имеем в виду наш «биологический оптимум», то для него создаются совершенно иные условия. Наше понимание оптимума не предполагает малой элиминации (и малой смертности), но предполагает, что она почти не имеет избирательного характера (малая интенсивность отбора). В этом случае плотность населения никогда не достигает предельного уровня, при котором начинается активная конкуренция за жизненные средства. Элиминация имеет вначале не косвенный, а прямой характер (потребляемость физическими факторами и хищниками подавляющей мощности). Поэтому она не приводит к снижению плодовитости, а, наоборот, статистические закономерности случайного переживания потомства поддерживают существование наиболее плодовитых линий, причем эта плодовитость при обеспеченности пищевым материалом полностью реализуется. Такой биологический оптимум возможен в двух случаях: во-первых, при свободном распространении в новой области, когда элиминация хищниками не велика, и во-вторых, даже при стационарном существовании в определенных условиях, когда прямая потребляемость очень велика, но не имеет избирательного характера.
Первый случай ограничивается во времени лишь периодом продолжающейся экспансии. Однако он наиболее интересен, так как сопровождается наиболее бурной мобилизацией и быстрой эволюцией по пути общего повышения организации (ароморфоз) или частного приспособления (алломорфоз).
Резкое ослабление остроты отбора (за исключением отбора на «общую жизнеспособность») ведет к быстрому вскрытию резервов изменчивости и повышению общей мутабильности. Изменчивость популяции достигает своего максимума в период ее расселения и увеличения численности на новых местах. Однако вскоре начинается либо размножение имевшихся уже хищников или паразитов, либо приспособление новых форм к хищничеству или паразитизму на новом иммигранте. Тогда элиминация приобретает избирательный характер и начинается очень быстрая (при большой эффективности отбора) эволюция в различных направлениях выработки мер защиты, частных приспособлений и т. д. Если же в завоеванной новой области действительно нет хищников, уничтожающих новую форму (как было, например, при первом завоевании суши членистоногими, а затем — позвоночными и вообще при всяком вторжении в еще необитаемые области новых водоемов или новых островов), то прогрессивное увеличение плотности населения ведет к развитию конкуренции, т. е. хотя и косвенной, но строго избирательной элиминации. В качестве примера можно сослаться на богатейшее видообразование рыб рода Haplochromis в озере Виктория, где не было активных хищников (Worthington, 1940), а также на видообразование бычков в оз. Байкал. Эффективный в этом случае отбор ведет к максимальной дифференциации при расхождении признаков и дальнейшей специализации, связанной со строжайшим распределением ролей (адаптивная радиация новых форм).
Во втором случае, когда прямая истребляемость велика и не допускает большого увеличения численности, но вместе с тем и не имеет избирательного характера (ввиду мощности стихийных сил неорганической или органической природы — климата или хищников подавляющей силы), повышение изменчивости приобретает длительный характер. Большая цифра гибели компенсируется высокой плодовитостью. Свободное накопление мутаций ведет к дегенеративным явлениям, к измельчанию, общему недоразвитию или преждевременному созреванию, в которых иногда и создается выход из создавшегося трудного положения (скрытая жизнь измельчавших форм). Это — путь катаморфоза, который может привести, при усиливающемся истреблении, и к «дегенеративному» вымиранию (Шмальгаузен, 1939а).
Ясно, что чем выше объем мобилизационного резерва, т. е. чем выше развит регуляторный аппарат организма, допускающий в широком масштабе скрытое мутирование и накопление нейтрализованных мутаций, тем выше потенциальная мобильность организма. Соответственно наиболее мобильны позвоночные животные, которые и достигали максимально высоких темпов эволюции при благоприятных для этого условиях.
Скорость вскрытия резервов и возможность их мобилизации очень хорошо видны на процессах одомашнения животных и введения в культуру различных используемых человеком растений. При условии ухода и охраны со стороны человека и главное подкармливания, у домашних животных потеряли свое значение почти все элиминирующие факторы. Естественный отбор если и не прекратился, то почти утратил свое влияние; остался лишь отбор на общую «жизнеспособность» в создаваемых человеком условиях. Поэтому должно было идти быстрое вскрытие резервов, которое затем и привело при участии искусственного отбора к быстрому породообразованию. В нем участвовали и новые мутации, возникавшие в особенности в благоприятствуемом направлении. Предел быстрого начального формообразования был, очевидно, продиктован объемом накопленного ранее резерва. Предел дальнейшей селекции диктуется физиологическими условиями «жизнеспособности» организма.
Ускорению мобилизации резервов и их обогащению способствовала также гибридизация, издавна применявшаяся человеком и приводившая к нарушению внутриклеточного «генного баланса» и к некоторому расшатыванию прочих регуляторных механизмов.
Мало специализированные исходные формы, жившие в благоприятных условиях и обладавшие большим запасом изменчивости, дали начало очень большому разнообразию пород — домашняя курица, собака, овца. Специализированные формы, жившие в более жестких условиях борьбы за существование и обладавшие соответственно меньшим запасом изменчивости, оказались менее мобильными и не дали такого пышного породообразования — домашняя утка и гусь (перелетные птицы с периодически более жестким отбором), кошка (специализированный хищник с острой конкуренцией; гибридизация, очевидно, не происходила), лошадь (специализированное травоядное с острой зимней элиминацией).
Из культивируемых растений скрытый запас изменчивости должен был быть выше у полиплоидов. Большое число сортов пшениц связано, очевидно, с их полиплоидностью. То же самое относится и к наиболее распространенному декоративному растению — культурным сортам роз. Впрочем, у растений общая картина сложнее, так как к этому присоединялись не только более свободная гибридизация и усиленное мутирование в условиях, создаваемых искусственными средствами культивирования, но также и различные другие факторы, в том числе соматические мутации, используемые методами вегетативного размножения и прививок. Все это способствовало еще более полному выявлению латентных свойств, чем это было возможно у животных.
Во всяком случае, и породообразование домашних животных и сортообразование культивируемых растений произошло с такой исключительной скоростью, очевидно, главным образом за счет накопленного ранее резерва изменчивости. Дальнейшая строго направленная селекция идет уже медленнее, хотя возможность выявления более мутабильных линий, а также гибридизации и влияния внешних воздействий позволяют и здесь рассчитывать на гораздо большие скорости и не ставит в этом смысле никаких ограничений (за исключением физиологически допустимых пределов).
в. Иммобилизация и специализация видов. Когда распространившаяся новая форма осела на новых местах или в новых экологических нишах и достигла известной плотности населения, начинает иногда сказываться постоянная или временная, периодическая или случайная нехватка жизненных средств. Тогда борьба за существование обостряется и в особенности возрастает активное соревнование особей за жизненные средства. В этом случае элиминация приобретает все более индивидуальный характер. Естественный отбор на общую жизнеспособность (при наиболее обеспеченных условиях существования) переходит в отбор на наибольшую организованность (выносливую конституцию) и приспособленность. Чем острее конкуренция, тем далее идет экологическое расхождение, тем более частный характер приобретает приспособленность организма, переходящая мало-помалу во все более узкую специализацию (теломорфоз).
Ведущая форма отбора теряет свое значение у максимально приспособленного организма и уступает место стабилизирующей форме. Влияние последней несколько различно в различных условиях борьбы за существование. Если лабильный организм не слишком специализирован и живет в разнообразной среде или если он даже специализирован, но живет в условиях постоянных и случайных изменений (непериодических колебаний) в факторах внешней среды, то стабилизирующий отбор (элиминация вредных мутаций и неблагоприятных модификационных уклонений от приспособленной нормы) ведет именно к стабилизации, т. е. к максимальной устойчивости формообразования в условиях изменчивой среды. Если же специализированный организм живет в однообразных и относительно постоянных условиях среды, то стабилизирующий отбор (т. е. элиминация всех — в данных условиях именно наследственных — уклонений от нормы) ведет к иммобилизации, т. е. к утере эволюционной пластичности. Иммобилизация есть результат острой элиминации всех мутаций, выражающихся в уклонениях от нормальной организации и ее нормальных функций. Она препятствует накоплению новых резервов изменчивости. Элиминируются, однако, именно проявляющиеся мутации, т. е. мутации, имеющие морфофизиологическое выражение. Поэтому иммобилизация достигает различного уровня у организмов, отличающихся по степени развития своего регуляторного аппарата.
Наиболее значительной должна быть иммобилизация гаплоидного организма, так как при отсутствии важнейших внутриклеточных регуляторных механизмов, связанных с диплоидно-стью, почти любая мутация должна иметь свое выражение и поэтому подлежит элиминации. Таким образом, гаплоидный организм не может иметь значительного скрытого мобилизационного резерва.
Редукция гаплоидного поколения у растений связана, вероятно, именно с его иммобильностью, т. е. эволюционным консерватизмом.
Так как мутации получают, во всяком случае, более значительное выражение при их переходе в гомозиготное состояние, то и гомозиготизация ведет к усиленной элиминации наследственных уклонений, и следовательно, к иммобилизации специализированных организмов. Все, что связано с родственным размножением, ведет поэтому к иммобилизации видов и утере эволюционной пластичности. Поэтому все формы более полной изоляции небольших популяций означают вместе с тем их иммобилизацию. В особенности, конечно, в тех случаях, когда мы имеем дело со специализированным организмом, живущим в весьма определенной и относительно константной среде и обладающим притом слабо развитой регуляторной системой (например, у животных с мозаичным развитием).
Консерватизм очень многих беспозвоночных связан с их специализацией и с жизнью в весьма однородной и постоянной среде. Это касается в особенности морских животных, живущих в защищенной обстановке, или глубоководных, а также малоподвижных и сидячих форм с малой потребляемостью (т. е. с преобладанием косвенной элиминации). Сидячие кишечнополостные, в том числе кораллы, многие двустворчатые и брюхоногие моллюски, частью и иглокожие, сидячие кольчецы, а в особенности брахиоподы отличаются удивительным консерватизмом на протяжении долгих геологических периодов. Этому способствовал и мозаичный характер развития высших червей, моллюсков и брахиопод. Исключение составляют головоногие моллюски — подвижные и активные хищники, обладающие хорошо развитым регуляторным аппаратом. Соответственно, они оказались в общем достаточно мобильными. Из хордовых животных консервативными оказались оболочники — специализированные организмы с мозаичным развитием, а наиболее мобильными — позвоночные, особенно высшие позвоночные, гораздо более универсальные по своей организации и обладающие наиболее развитой системой морфогенетических корреляций регу-ляторного характера и соответственно, очевидно, значительным запасом наследственной изменчивости,
Шпет и Иванько (1940) сделали попытку сопоставить изменчивость представителей консервативных («древних») групп насекомых (Odonata, Blattoidea, Ephemeroptera) и изменчивость представителей прогрессивных групп (Diptera, Hymenoptera, Lepidop-tera). Получились биометрически ясные различия в пользу последних.
В практике разведения домашних животных и культивируемых растений мы имеем дело также со специализацией и иммобилизацией пород и сортов. Благоприятные условия размножения животных и культивируемых растений при неограниченных жизненных средствах и защите со стороны человека вели вначале ко вскрытию резервов изменчивости и их мобилизации. Это привело при применении искусственного отбора (в его «ведущей» форме) к быстрому породо- и сортообразованию. Однако дальнейшая, более строгая, селекция шла по пути более узкой и притом неограниченно далеко идущей «специализации» требований (отбор мясных, шерстных и т. д. пород), что привело к выработке стандартов и ко все более интенсивной браковке уклонений от этих стандартов. Это можно рассматривать как стабилизирующую форму искусственного отбора.
Однако искусственная стабилизация ведется теперь и в специально выработанных (пусть даже оптимальных) стандартных условиях развития и содержания. Следовательно, стабилизирующий отбор происходит в относительно весьма постоянных условиях внешней среды. Поэтому почти все уклонения имеют наследственный характер. Элиминация наследственных уклонений означает тогда именно иммобилизацию, т. е. утрату пластичности. Той же иммобилизации содействует, конечно, и внутрипородное родственное разведение. Впрочем, у домашних птиц и млекопитающих, обладающих высоко развитой системой морфогенетических регуляций, иммобилизация не может быть полной, так как скрытое мутирование ускользает от контроля человека.
То же самое происходит и с лабораторными и любыми экспериментальными культурами. Если они вначале ставятся в оптимальные условия существования и размножения, то имеет место быстрое вскрытие резервов — увеличение изменчивости. В дальнейшем происходит выделение известных линий и их стабилизация, а в условиях родственного разведения в выработанных стандартных условиях — их иммобилизация. В особенности это касается культур животных с мозаичным развитием и слабо развитой системой регуляций, как, например, дрозофилы.
Свеже поставленные культуры дрозофилы, взятой из природных популяций, показывают всегда вскрытие резервов (более значительных, если это большая популяция, менее значительных, если это небольшие изолированные популяции, и в особенности малых, если популяции жили в постоянных, ограниченных и суровых условиях острой элиминации). Однако лабораторная линия «Флорида», прошедшая уже в лабораториях огромное число поколений, из которых систематически элиминировалась каждая замеченная мутация, прошла таким образом путь жесткой стабилизации.
Так как эти культуры ведутся в стандартных условиях постоянной температуры (25°), постоянной влажности и стандартной кормовой смеси, то «стабилизирующий» искусственный отбор привел затем к неизбежной «иммобилизации». Лабораторная линия «Флорида» естественно теряет свою пластичность. По Р. Л. Берг, она так же мало мутабильна, как и вполне изолированные небольшие природные популяции дрозофилы. По мере дальнейшей культуры в стандартных условиях можно ожидать лишь усиливающегося снижения мутабильности.
10. КОНТРОЛЬ И РЕГУЛЯЦИЯ В ЭВОЛЮЦИИ
А. ЭВОЛЮЦИЯ КАК РЕГУЛИРУЕМЫЙ ПРОЦЕСС
Эволюция, т. е. процесс исторического развития организмов, протекает закономерно. Основные закономерности этого процесса были вскрыты еще Ч. Дарвином. Они выражаются в непрерывном приспособлении организмов во всех своих функциях и всех чертах своей организации к той среде, в которой они обитают и именно к тем факторам среды, с которыми они сталкиваются в своей жизнедеятельности. Это приспособление достигает иногда удивительного совершенства и исключительной тонкости и вместе с тем оно всегда совпадает именно с теми условиями, в которых протекает жизненный цикл данного организма. Если в течение жизненного цикла происходит закономерная смена среды обитания (например, переход от водной жизни к наземной, как у амфибий и у некоторых насекомых, или от скрытой жизни в земле или в растительных тканях к открытой жизни, как у многих жуков), то это всегда сопровождается соответствующей перестройкой организации. Если у организма имеются ненужные ему органы и приспособления, это указывает на исторический выход из другой среды и всегда ведет к постепенной редукции этих органов и приспособлений (рудиментарные органы). Такое точное приспособление организма к внешней среде не было бы возможно, если бы не существовало механизма, контролирующего и регулирующего исторический ход этого приспособления. Этот механизм управляет, очевидно, процессом эволюции организмов в связи с факторами внешней среды. Должна быть органическая связь между организмом и средой не только в его жизни (что и без того ясно), но и в ходе их исторических преобразований. Такая связь была найдена Ч. Дарвином в виде борьбы всех живых существ за свое существование в конкретной жизненной обстановке. Борьба за существование в условиях конкуренции в овладении жизненными средствами ведет к переживанию наиболее приспособленных особей любого вида организмов и их преимущественному размножению. В этом и состоит естественный отбор как основная часть регулирующего механизма эволюции. Теория естественного отбора полностью оправдала себя в течение дальнейшего развития биологических знаний. Она, однако, не дает полного объяснения всего эволюционного процесса.
Естественный отбор — это регулирующий механизм эволюции, но не ее исходная материальная основа. Ч. Дарвин указал и на эту основу — изменчивость и наследственность. Однако закономерности изменчивости и наследственности, несмотря на огромный эмпирический материал, собранный Дарвином, оставались в то время еще совершенно неизвестными. Только текущее столетие пролило свет на эти проблемы. Благодаря блестяшчм экспериментальным исследованиям, берущим свое начало от классических опытов Менделя, мы можем в настоящее время сказать, что в основном механизм изменчивости и наследственности уже вскрыт.
Таким образом, в настоящее время хорошо изучена материальная база эволюционного процесса и известен механизм его регуляции. Это — начальное и конечное звенья в элементарном цикле эволюционных преобразований. Роль этих звеньев достаточно ясна и легко поддается количественному учету. Для этого были введены понятия мутационного «давления» (т. е. скорости или частоты возникновения наследственных изменений) и «давления» естественного отбора (т. е. его эффективности в известных условиях). Эти понятия легли в основу генетической теории естественного отбора (Fisher, 1930). Вместе с экспериментальными исследованиями популяционноп генетики они положили начало точному анализу факторов эволюции. Несмотря на большое значение уже достигнутых результатов, такие исследования не вскрывают всего механизма эволюции и не дают полного объяснения его закономерностей.
При таком подходе в тени остается индивидуальное развитие организмов, ведущее к реализации фенотипа. Так как именно фенотипы являются активными носителями жизни и объектами естественного отбора, то ход индивидуального развития не может не иметь значения эволюции. И, наконец, самое главное, в генетической теории естественного отбора не видно организма как такового с его активной борьбой за свою жизнь. Понятие борьбы за существование, лежащее в основе теории Дарвина, совершенно выпало. Естественный отбор выступает как внешний фактор, а сам организм — как пассивный объект, с которым оперирует естественный отбор. Это не является верным отражением действительных соотношений. В оправдание установившейся в генетике точки зрения следует сказать, что дарвиновское понятие борьбы за существование сформулировано столь расплывчато, что с ним действительно трудно оперировать. Это видели многие критики и особенно К. А. Тимирязев, который предложил вообще заменить его понятием элиминация. Последняя поддается точному учету. Однако элиминация является лишь обратной стороной естественного отбора и в таком виде остается внешним фактором по отношению к организму.
Нужны, очевидно, новые понятия, во-первых, отражающие активную роль организма не только в своих жизненных проявлениях, но и в эволюции и, во-вторых, хорошо ограниченные и доступные точному исследованию. Возможно, что такие понятия удастся сформулировать, если подойти к эволюции с новыми представлениями об устройстве регулируемых механизмов вообще, т. е. с точки зрения кибернетики (Ashby, 1956) (включающей теорию информации). Как уже пояснено, любой автоматически регулируемый механизм составляется из двух частей — регулируемого объекта и регулятора, которые соединены между собой линиями связи, передающими управляющие сигналы от регулятора и обратную информацию о состоянии объекта к регулятору.
В процессе эволюции органического мира управляемым объектом является вид в целом. Однако вид организмов обычно разбит на подчиненные единицы, ведущие до известной степени самостоятельное существование и обнаруживающие нередко явные признаки приспособления к местным условиям. Наименьшей из таких единиц является популяция особей. Поэтому, рассматривая лишь элементарные основы эволюции, мы должны исходить из популяции как наименьшей эволюирующей единицы. Направление эволюционного процесса определяется, очевидно, внешней средой. На это ясно указывает точная приспособленность организмов к условиям их существования и при том не только к климатическим, эдафическим и другим факторам неорганического окружения, но и к тем живым существам, с которыми они сталкиваются в этой среде и с которыми они связаны теснейшими взаимоотношениями (и прежде всего пищевыми связями). С другой стороны, нельзя преуменьшать и роли внутренних факторов. На их значение указывает точная слаженность организации, взаимоприспособленность отдельных частей, органов и тканей, согласованность их функций. Наконец, и в выборе местообитания, и в столкновениях с другими организмами, и в пищевых взаимоотношениях активную роль играют и сами особи данной популяции.
Таким образом ясно, что регулирующий механизм кроется в некоторых формах взаимодействия между организмом и внешней средой. Этот механизм и был вскрыт Ч. Дарвином. Его действие проявляется в различных формах борьбы за существование, ведущей через дифференциальное переживание и размножение к естественному отбору наиболее приспособленных особей. Борьба за существование проявляется в теснейшем взаимодействии организма и среды (включающей все другие организмы, совместно с ним обитающие и образующие вместе с ними единое «сообщество» — биоценоз). Так как мы не можем отрицать значение факторов неорганической среды, в которой живет и размножается данный организм, мы будем говорить о биогеоценозе (Сукачев, 1945) как о регулирующем аппарате эволюции. Любая популяция особей определенного вида, входящая в состав известного биогеоценоза, является его органической, необходимой составной частью. Таким образом, в данном случае обнаруживается самая интимная связь между управляемым объектом — популяцией и регулятором — биогеоценозом. Такая тесная связь наблюдается нередко и в конструируемых нами механизмах, где управляемый объект является как бы частью регулятора или, наоборот, регулятор входит как часть в состав управляемой машины. Существование тесной взаимосвязи и является условием, допускающим возможность контроля и управления эволюционным процессом. Если биогеоценоз в целом играет роль регулятора эволюционного процесса, то он обязательно должен быть обеспечен «информацией» о состоянии популяции (по линии «обратной» связи), должен включать в себя специфический механизм преобразования этой информации в управляющие сигналы и средства передачи последних на популяцию. Таким образом, кроме механизма преобразования требуется наличие каналов связи для передачи информации в двух направлениях — от популяции к биогеоценозу и от биогеоценоза к популяции. Так как изменение популяции, как элементарный эволюционный процесс, всегда сопровождается наследственным изменением ее особей, то управляющие сигналы от биогеоценоза к популяции должны каким-то образом включать возможность изменения ее наследственной структуры. Последнее может произойти только в процессе преобразования информации в самом биогеоценозе (т. е. в «регуляторе»). Так как первичные эволюционные изменения возможны только в популяции (или в поколениях особей, но не в отдельных особях), то наиболее простым изменением является хотя бы небольшое изменение в генетическом составе популяции, т. е. в соотношении числа особей с разной наследственной характеристикой (генотипов). Информация о таких изменениях популяции может быть сообщена через наследственный аппарат ее особей и передана особям следующего поколения при посредстве половых клеток или иных средств воспроизведения. Такой аппарат действительно имеется и несомненно он полностью обеспечивает надежную связь популяции с регулирующим механизмом биогеоценоза и дальнейшую передачу информации от одного поколения особей к следующему. Имеются и средства передачи обратной информации от популяции к биогеоценозу.
Популяция, несомненно, активно воздействует на биогеоценоз, хотя бы через потребление пищевых материалов и накопление продуктов своей жизнедеятельности. В известных условиях популяция может внести значительные изменения в строение биогеоценоза. Таким образом, имеются и каналы обратной связи.
Однако нет прямой связи между наследственной информацией по первому каналу (от биогеоценоза) и обратной информацией по второму каналу (от популяции к биогеоценозу). Здесь непосредственная связь как будто прерывается, так как обе линии связи находятся на разных уровнях. Наследственная информация передается на внутриклеточном (молекулярном) уровне организации, а обратная информация только на уровне организации целой особи.
Переход от одной линии связи к другой совершается здесь (как обычно и в технике) через посредство довольно сложного механизма преобразования. Наследственная информация преобразуется в процессах индивидуального развития в средства передачи обратной информации, именно — в фенотип особи, являющейся реальным носителем жизни и активным участником наступления на жизненные ресурсы биогеоценоза («борьбы за существование»). В биогеоценозе через естественный отбор и процессы размножения происходит новое преобразование этой информации в наследственную с переходом от уровня организации особи (в фенотипах) на уровень организации клетки (половые клетки, зиготы) . Этим и замыкается полный круг преобразований в элементарном цикле эволюционного процесса (табл. 3).

Таким образом, основные требования, предъявляемые к конструкции автоматически регулируемого устройства, в данном случае оказываются выполненными, и мы можем попытаться осветить более подробно работу отдельных звеньев этого механизма. Это необходимо сделать для того, чтобы представить себе яснее, может ли новая точка зрения внести что-либо новое в понимание эволюционного процесса, выдвигает ли она новые вопросы, освещает ли она наиболее ответственные звенья этого процесса. Не позволит ли она сформулировать основные понятия таким образом, чтобы внести полную ясность и четкость в понимание механизма эволюции и сделать все его звенья доступными для точного количественного анализа.
Б. НАСЛЕДСТВЕННАЯ ИНФОРМАЦИЯ И СРЕДСТВА ЕЕ ПЕРЕДАЧИ
Наследственная информация передается от материнских особей к дочерним особям, т. е. от предыдущей генерации к последующей генерации той же популяции. В предшествующем изложении мы говорили также о передаче информации от биогеоценоза к популяции. Нет ли здесь неувязки? Нет, неувязки здесь нет, но нужно учесть, что наследственная информация передается от предыдущей генерации к последующей только после ее преобразования в биогеоценозе. Схема популяции входит в состав биогеоценоза в качестве его неотъемлемой части. Механизм контроля и преобразования информации заключается в самом биогеоценозе, именно во взаимодействии особей данной популяции с другими членами и факторами биогеоценоза. Преобразование совершается через естественный отбор, т. е. при преимущественном размножении одних особей с их наследственными механизмами и элиминации других. Таким образом, под влиянием естественного отбора в самом биогеоценозе происходит изменение состава популяции, меняется ее наследственная характеристика. Во всей популяции в целом создаются новые соотношения по меньшей мере в численности различных генотипов. Механизм наследственной передачи лежит, следовательно, в отдельных особях, а изменение в биогеоценозе касается не отдельных особей, а всей популяции в целом. Информация об этих изменениях передается все же через посредство размножения апробированных особей, потомство которых вливается в ту же популяцию. Таким образом меняется популяция, но информация об этом изменении (происшедшем в биогеоценозе) передается через размножение особей.
Средством передачи наследственной информации являются либо половые клетки, либо другие пропагационные клетки, образующие зачатки нового организма при вегетативном размножении. Во всяком случае такая передача осуществляется только через посредство клеток. Механизм этой передачи хорошо известен, он совпадает с механизмом клеточных делений, при котором полностью обеспечивается равномерное распределение основных ядерных субстанций в виде определенного набора хромосом. Каждая клетка получает полный набор этих хромосом. Изучение строения последних с помощью двух принципиально различных методов — гибридологического и цитологического — показало их сложный состав из большого числа наследственных единиц, располагающихся в ряд по всей длине хромосомы. Гибридологической единицей наследственности является ген. Изучение химического состава хромосом показало, что важнейшей их составной частью является дезоксирибонуклеиновая кислота (ДНК). Молекула последней имеет вид очень длинной парной цепочки, состоящей из чередующихся фосфатных и сахарных групп, связанных парами пурино-вых и пиридиновых оснований, образующих как бы перекладины веревочной лестницы (Watson, 1953). Строение этих гигантских молекул обеспечивает как высокую индивидуальность, так и значительную их стойкость. Оно дает также возможность продольного расщепления молекулы с последующим восстановлением каждого партнера. Очевидно, такое расщепление и восстановление и лежит в основе расщепления хромосом и их генов при клеточных делениях. Во всяком случае это гарантирует возможность точной передачи наследственной информации от клетки к клетке.
Вся полнота информации о наследственных свойствах зиготы и развивающейся из нее особи обеспечивается определенной последовательностью и положением двух возможных пар оснований (аденин с тимином или гуанин с цитозином) в каждой молекуле. При большом числе звеньев, исчисляемом тысячами, комбинированное расположение четырех качественно различных знаков (оснований) гарантирует возможность записи наследственных структур любой сложности в виде условного «кода». Наследственная информация передается, следовательно, в закодированном виде и при том вся единовременно в пространственной связи, а не во временной последовательности. При расшифровке этого кода в процессах индивидуального развития информация развертывается также в пространстве, хотя ее преобразование совершается несомненно во временной последовательности. Химической единицей нашего кода является, как видно, пара оснований, которая может принять форму лишь четырех знаков (аденин с тимином, гуанин с цитозином, или в противоположном положении — тимин с аденином и цитозин с гуанином). Чередование этих знаков может обеспечить передачу любой информации (как телеграф Морзе с таким же числом знаков). Гибридологической единицей наследственности является, однако, более сложное образование — ген как выражение системных связей в хромосомах (об этом см. дальше в разделе Ж).
В. ПРЕОБРАЗОВАНИЕ ИНФОРМАЦИИ В ИНДИВИДУАЛЬНОМ РАЗВИТИИ
Преобразование информации представляет всегда закономерный процесс, полностью определяемый устройством той системы, в которой совершается это преобразование. Наследственная информация, заключающаяся в зиготе или в другой пропагационной клетке, преобразуется затем в процессах индивидуального развития. Характер преобразования определяется организацией зиготы и поэтапно развивающейся из нее особи. Это преобразование представляет собой очень сложный процесс (полное рассмотрение которого выходит за пределы поставленной нами задачи). Он строится на биохимической основе внутриклеточного обмена, регулируемого ядерными субстанциями хромосом. Однако в эти процессы, естественно, вовлекаются и плазменные субстанции, взаимодействующие с ядерными продуктами в процессах этого обмена (табл. 4). Первоначальные дифференцировки зиготы и дробящегося яйца определяются именно организацией плазмы, детерминированной еще в яичнике материнского организма. Дальнейшая дифференцировка идет иногда при заметном участии внешних факторов, но определяется все же взаимодействием частей развивающегося организма. Такие взаимодействия частей осуществляются в «индукционных», «формообразовательных» и «корреляционных» системах, которые имеют более или менее ярко выраженный регуляторный характер (гомеостат). Внешние факторы не имеют специфического формообразовательного значения. Они оказываются лишь в роли освобождающих причин, определяющих время и иногда место наступления известной реакции, и в лучшем случае служат для включения «стрелочного» механизма, направляющего развитие по одному или другому из существующих уже, т. е. полностью детерминированных, путей («каналов») развития. На более поздних стадиях развития регуляция формообразования переходит к эндокринной системе и к самой функции отдельных органов, связанных через посредство кровеносной и, в особенности, — нервной системы. В функциональных зависимостях вновь ясно выражается связь с внешней средой и влияние факторов среды на формообразование. Однако это влияние всегда является косвенным и не определяет специфики морфологических процессов. Таким образом, если некоторые функции контроля и выполняются внешней средой, то все же специфические ответы на факторы этой среды определяются унаследованными нормами реакции данного организма. Только поэтому фенотип особи и является полным и точным (для данных условий среды) выражением генотипа. Регуляторные функции выполняются всегда унаследованным механизмом развития самого организма. Поэтому всякое развитие организма (особи) есть по меньшей мере авторегуляция. В высших формах индивидуального развития зависимость от факторов внешней среды теряет в еще большей мере свое формообразовательное значение и авторегуляция переходит в более или менее ярко выраженное автономное развитие. Как и все формы преобразования информации, индивидуальное развитие определяется в своей специфике только организацией самого преобразующего механизма. Во всех случаях формообразования оно индуцируется очень незначительным влиянием, небольшой разницей в обмене, незначительным количеством специфических продуктов индуктора или эндокринной железы, ничтожным поступлением энергии из внешней среды и реализуется в полной мере при малейшем превышении порогового уровня реактивности материала. Реакция ни в какой мере не пропорциональна формообразовательному воздействию. Она осуществляется на известном его уровне сразу и в полном масштабе, определяемом унаследованной «нормой». Обычно она выражается в клеточных делениях, перемещениях и дифференцировке больших клеточных масс. Реакция имеет по существу «взрывной» характер. Она подготовлена заранее в виде известного запаса энергии внутри реагирующего материала. Такой характер индивидуального развития, т. е. преобразования наследственной информации, напоминает обычный метод преобразования информации с использованием специфических усилителей в регулирующих механизмах автоматического управления.
Наследственный материал хромосом является средством передачи унаследованной структуры (т. е. передачи закодированной информации по первому каналу связи) от материнского организма через половую клетку и зиготу ко всем клеткам дочернего организма. Хромосомы являются, однако, не только средством механической передачи химических веществ определенной унаследованной структуры от клетки к клетке. Хромосомы являются прежде всего активными регуляторами внутриклеточного обмена веществ и контролируют через это все процессы индивидуального развития. Наследственная информация реализуется через эти процессы в виде развивающейся особи — фенотипа. Фенотип является внешним выражением генотипа и вместе с тем активным руководителем внеклеточного обмена веществ между организмом и внешней средой. Он контролирует всю его жизнедеятельность. И вместе с тем фенотип является средством обратной информации от организма к внешней среде (биогеоценозу). Эта передача производится совсем иными средствами, чем передача наследственной информации от материнского организма к дочернему. И все же она находится в органической связи с этой передачей — фенотип вполне закономерно отображает генотип, но только в совершенно иной форме. В индивидуальном развитии происходит преобразование скрытой, наследственной информации (записанной в виде условного кода в хромосомах) в явную информацию об унаследованных нормах реакции. Это преобразование происходит во взаимодействии с факторами внешней среды, которые являются необходимыми условиями и, иногда, освобождающими факторами этого преобразования. Вся его качественная специфика в данных условиях определяется, однако, унаследованными нормами реакций, т. е. генотипом. Поэтому фенотип является вполне реальным выразителем генотипа в известных условиях развития. Фенотип есть преобразованный генотип и если последний закодирован в тончайшей структуре хромосом и управляет внутриклеточным обменом (т. е. жизнедеятельностью клетки), то первый обнаруживается во всех признаках целого организма (особи) и проявляется в специфических формах его жизнедеятельности. Наследственная структура хромосом является средством межклеточной связи внутри организма (от зиготы к первичной половой клетке через многочисленные клеточные деления) и регулятором внутриклеточного обмена веществ. Фенотип является средством связи между особью и внешней средой и регулятором обмена веществ между особью и этой средой. В обоих случаях эти структуры ответственны за основные жизненные проявления, но только хромосомы — в масштабе клетки, а фенотип — в масштабе целой особи. Фенотип также «записан» условными символами, как и генотип, но только это совершенно иные символы, так как они несут хотя и сходную функцию, но на различных уровнях организации в разных условиях среды.

Наследственная информация, передающаяся на молекулярном уровне организации и реализующаяся в процессах внутриклеточного обмена, преобразуется в обратную информацию, передаваемую на уровне организации особи и реализующуюся в процессах жизнедеятельности, т. е. обмене веществ целой особи. Весь смысл индивидуального развития состоит в преобразовании наследственной информации в систему жизненных связей организма с внешней средой. Эти связи устанавливаются посредством признаков фенотипа, которые одновременно являются и средствами, позволяющими осуществить контроль этой организации со стороны факторов биогеоценоза. Поэтому индивидуальное развитие организма, связанное с реализацией (в известных условиях) наследственной структуры в фенотипе развивающейся, а затем и зрелой особи, представляет типичный пример преобразования информации с переходом от одного кода к другому.
Г. ПЕРЕДАЧА ОБРАТНОЙ ИНФОРМАЦИИ ОТ ПОПУЛЯЦИИ К БИОГЕОЦЕНОЗУ
Средством передачи обратной информации является фенотип и именно специфические формы активности каждой особи, как разнообразные выражения ее жизнедеятельности в определенной обстановке— в биогеоценозе.
Основным проявлением активности является добывание жизненных средств и прежде всего потребление пищевых материалов (пищевые материалы используются на развитие, рост, на поддержание жизни и на затраты энергии при всех проявлениях жизнедеятельности). Вторым проявлением активности является деятельность, связанная с размножением — половая активность и воспроизведение потомства. Третьим проявлением жизненной активности является деятельность, связанная с защитой своей жизни и жизни потомства. Обратная информация передается, таким образом, через весь фенотип в целом. Это не значит, что эта информация неразложима. Как геном при всей своей целостности разложим на единицы наследственной информации — гены и даже еще далее — на простейшие химические единицы (пары оснований), так и целостный фенотип разложим на многочисленные компоненты, являющиеся также своего рода символами, отображающими наследственные свойства особи. Связь между этими символами или признаками является весьма тесной. И все же некоторые признаки могут быть разделены и перекомбинированы или изменены в результате соответствующих изменений в наследственных единицах — генах. Это касается прежде всего таких поверхностных признаков, как окраска, рисунок, структура покровов, форма и окраска волос и многие другие. Отдельные признаки внутренней организации также могут быть изменены и это приводит к изменениям в обмене, в функциях отдельных органов, в поведении. Чаще всего, однако, такие изменения затрагивают целые взаимосвязанные системы, регулирующие основные жизненные проявления, и тогда их результат сказывается в изменении общей выносливости организма по отношению к внешним факторам (климату), общей активности, жизнедеятельности, плодовитости.
Взаимосвязанность большинства признаков фенотипа является условием жизнеспособности, устойчивости и активности особи и вместе с тем условием, обеспечивающим высокие качества передачи информации. В каждой особи признаки фенотипа обнаруживают высокую степень «сцепления». Они настолько связаны, что образуют в сущности одну целостную систему. Каждая особь глубоко индивидуальна и является надежным носителем высококачественной информации. Последняя имеет и в этом случае (как и в геноме) «картинный» характер, она развертывается не во временной последовательности, а в пространстве. Хотя она передается и во времени, но при этом различные ее комплексы выявляются все же сразу. Во времени меняется и фенотип и, следовательно, сама информация (возрастные изменения), но и в этом случае меняется одновременно вся «картина». Обратная информация от популяции передается через посредство не одной особи, а всех особей данной популяции. Особь является лишь самостоятельной единицей этой информации. Поэтому вся информация оказывается гораздо более сложной. При максимальном разнообразии особей, когда каждый фенотип (как и генотип) неповторим, количество информации является максимальным. Во вполне однородной популяции количество информации падает до нуля и регуляция ее состава оказывается невозможной.
Особи являются носителями равновероятных сообщений, если фенотипы равномерно распределены в популяции. Они могут, однако, группироваться по некоторым признакам фенотипа в подчиненные единицы разной численности (полиморфизм). Такие группы получают свою качественную характеристику, могут получить положительную оценку в известных условиях и дать начало новой самостоятельной популяции и новому таксономическому подразделению. Особи одной популяции являются, как правило, изолированными носителями сходной информации. Этим гарантируется повторность и, следовательно, надежность информации по наиболее существенным признакам организации. Одновременно по второстепенным признакам этой повторности нет, так как в этом отношении проявляются большие различия между особями. Многообразие особей при их изолированности делает состав популяции весьма подвижным — в процессе эволюции он легко меняется (через контроль и процесс преобразования информации в биогеоценозе).
Однако не всегда особи одной популяции являются вполне изолированными единицами. В некоторых случаях между особями данной популяции или ее части устанавливается более тесная связь. Выделяются временные группы особей, называемые стадами, стаями, косяками или, при более тесной и постоянной связи по местообитанию — колониями. Такое объединение особей ведет к улучшению качества информации по меньшей мере в результате увеличения помехоустойчивости (см. далее). Попарное объединение особей разного пола ведет к более надежной передаче информации следующему поколению (по первому каналу), но связано и с некоторым повышением помехоустойчивости в передаче обратной информации.
Передача обратной информации от популяции к биогеоценозу осуществляется, как сказано, через посредство отдельных особей и выражается в различных формах жизненной активности. Прежде всего эта активность проявляется в добывании жизненных средств. Она строго специфична, так как особи одного вида потребляют совершенно определенные неорганические и органические вещества. Она до некоторой степени индивидуализирована и выражает потребности, особенности, вкусы и привычки данной особи. Животные, как правило, более активны и истребляют обычно большое количество других организмов. Однако даже наиболее пассивные виды живых существ оказывают значительное влияние на внешнюю среду и при отсутствии сопротивления своей деятельности быстро могли бы изменить весь состав биогеоценоза (что иногда и наблюдается). Всей своей жизнедеятельностью особи любого вида организмов воздействуют на биогеоценоз и тем самым «информируют» его о состоянии своей популяции. Свойства особи (фенотипа) данной популяции проявляются в формах ее активности (жизнедеятельности). Однако они бывают отмечены и внешними признаками, характеризующими особей данного вида и также в значительной мере индивидуализированными. Эти внешние признаки играют также известную роль как средства информации, передаваемой от особи к особи той же популяции, а иногда и к особям других видов. Эта информация входит в понятие обратной информации и подлежит такому же контролю и преобразованию в биогеоценозе.
Наглядным примером хорошо видимой обратной информации и ее значения в контроле и регуляции эволюционного процесса является информация, передаваемая другим особям того же вида (как элементам того же биогеоценоза) через посредство внешних сигналов — видовых признаков окраски, формы, поведения, звуков, запахов и т. д. Через посредство этих признаков особи одного вида связываются между собой иногда для совместных форм жизнедеятельности и защиты (стадный образ жизни многих млекопитающих, стаи птиц, косяки рыб) и, во всяком случае, они являются сигналами безопасности, способствующими спокойным проявлениям нормальной жизнедеятельности.
Еще более заметно значение передачи тех же или подобных сигналов особям другого пола, как средства, способствующего спариванию, т. е. проявлению половой активности. Большое значение этих сигналов приводит иногда к очень резкому их выражению и даже к кажущейся гипертрофии. Таковы ярко выраженные вторично-половые признаки многих животных. Все они способствуют спариванию, т. е. оставлению потомства. Все они являются внешними сигналами, отмечающими активность особей. Во всех таких признаках проявляются и формы индивидуального состязания, особенно активного часто между самцами. Они ведут к естественному отбору (половому отбору) наиболее эффективных средств информации через контроль данной формы жизнедеятельности (т. е. спаривание и оставление потомства).
Все эти формы информации являются частными, но весьма наглядными примерами, показывающими значение обратной информации для контроля и регулирования эволюционного процесса.
Д. КОНТРОЛЬ ФЕНОТИПОВ В БИОГЕОЦЕНОЗЕ
Популяция воздействует на биогеоценоз через посредство активной деятельности ее особей. Это мы рассматриваем как обратную информацию о состоянии популяции.
Активная деятельность особей контролируется внешними по отношению к популяции факторами биогеоценоза, именно, с одной стороны, организмами, для которых особи данной популяции являются пищевым материалом и, с другой стороны, наличием пищевых материалов и других жизненных средств, необходимых для поддержания собственного существования и для оставления потомства.
Уничтожающие факторы имеют всегда случайный характер и они не входят в механизм контроля как необходимый его элемент. Более того, массовое уничтожение особей случайными внешними факторами может иметь лишь отрицательное значение в жизни и эволюции популяции. Вред прямого истребления снижается по мере уменьшения их ценности (и доступности) как кормового материала и, однако, даже полная несъедобность не может стать для них фактором прогрессивной эволюции. Фактором прогрессивной эволюции может быть лишь активная роль популяции в поддержании своей жизни и размножения и в защите своей жизни и жизни потомства. В этой активности выражаются наследственные свойства фенотипов и их способность добывать необходимые жизненные средства.
Поэтому, учитывая контролирующее значение жизненных ресурсов внешней среды, мы все же выдвигаем значение впутренних соотношений в самой популяции и в особенности форм жизнедеятельности и уровня активности отдельных ее особей.
Как уже неоднократно отмечалось, обратная информация от популяции к биогеоценозу передается именно через посредство форм активности ее особей (фенотипов), из которых важнейшей является добывание жизненных средств. Контролируется и ограничивается эта активность в биогеоценозе наличными запасами пищевых материалов и их доступностью, т. е. затратой энергии на их добывание. Максимум использования при минимальной затрате энергия является источником сил и сопротивляемости организма. Наиболее активные особи нормально развиваются и оставляют потомство. Менее успевающие и потому менее устойчивые особи гибнут в борьбе с хищниками, болезнями или климатическими невзгодами. Результатом жизненного цикла особи может быть либо поддержание своей жизни вплоть до оставления потомства, либо преждевременная гибель без оставления потомства. Так как результат определяется при наличии ограниченных запасов жизненных средств в основном индивидуальными качествами особей — формами и степенью их активности, то их «оценка» является сравнительной. Такую сравнительную оценку можно назвать состязанием, соревнованием или конкуренцией (если она достигает известной остроты при нехватке жизненных средств). Решающими в этом состязании являются именно индивидуальные качества особей данной популяции. Таким образом, это состязание является внутригрупповым. Понятно, что при отсутствии различий между особями, т. е. во вполне однородной популяции, такое состязание не может привести ни к какому результату. Состязание происходит между разными особями внутри популяции. Результаты состязания определяются, конечно, не только составом популяции, но и запасами жизненных средств во внешней среде, т. е. взаимодействием внешних и внутренних факторов. Таким образом, механизм контроля осуществляется силами биогеоценоза (куда входит и популяция). Основным, но не единственным критерием служит при этом соотношение между активностью каждой особи и наличными запасами жизненных средств в биогеоценозе.
Объектом контроля является фенотип особи, а средством контроля — те факторы среды, с которыми особь нормально сталкивается в своей жизнедеятельности (в первую очередь — жизненные средства). Каждая особь соприкасается со многими внешними факторами и испытание ее качеств происходит многократно в течение всего жизненного цикла, т. е. на всех стадиях ее развития. При этом происходит не только последовательная смена внутренних факторов, характеризующая онтогенез особи и его потребности, но также и смена факторов внешней среды с ее запасами различных жизненных средств. Иногда это — довольно резкая смена всей жизненной обстановки, связанная с коренным изменением форм активности (эмбриональный период, личиночный, возрастные изменения, смена форм дыхания и питания, сезонные и локальные изменения, активный переход из одной среды в другую и т. д.).
Все закономерные изменения во внешних факторах играют свою роль в контролировании жизнедеятельности отдельных особей и всей популяции в целом. Метод оценки особей таким образом весьма сложен и совершенен. При этом используются все факторы среды, связанные так или иначе с жизнью особи. Все необходимые факторы для поддержания жизни мы называли суммарно жизненными средствами. В этой сложной обстановке соревнования решающее значение приобретают индивидуальные качества особей. Очевидно, мы и эти качества особей можем суммарно обозначить как способность извлекать и использовать жизненные средства. Хотя это и является основным в оценке фенотипа, все же нельзя не учитывать и другого — способности уклоняться от неблагоприятных влияний, которые могут преждевременно прервать жизнь особи. Учитывая все свойства особей (фенотипов), которые имеют решающее значение при сравнительной их оценке в биогеоценозе, мы говорили о формах жизнедеятельности, или формах активности отдельных особей. Эти качества, несомненно, доступны для экспериментального исследования и количественного учета.
Состязание особей в способности добывать и использовать жизненные средства является основой теории Дарвина. Контрольный механизм биогеоценоза представляет собой не что иное как «борьбу за существование» Дарвина, которая связана с естественным отбором.
Мы, однако, сознательно несколько упростили здесь представление о борьбе за существование. Мы ограничились рассмотрением состязания в формах жизнедеятельности и притом главным образом в овладении жизненными средствами и оставили на втором плане состязание в средствах борьбы с неблагоприятными внешними влияниями климата, с болезнями, хищниками и т. п. Хотя и эти факторы играют известную роль контролирующего механизма и решающее значение здесь также имеют различные формы активности организма, мы их рассмотрим в дальнейшем отдельно (см. раздел «Значение помех...»), так как результаты их влияния существенно отличны. Несомненно, что основное ведущее значение имеет контроль активности особей в овладении жизненными средствами.
Таким образом, во всей бесконечно сложной и поэтому трудно анализируемой «борьбе за существование» большое значение имеет лишь контроль фактического использования пищевых материалов биогеоценоза. Контроль этот всегда оказывается многостепенным. Предварительным результатом положительной оценки оказывается переживание, спаривание и размножение данной особи. Дальнейшая оценка осуществляется через переживание, спаривание и размножение фактически оставленного потомства данной пары и т. д.
Контролирующий механизм биогеоценоза осуществляет свою функцию через посредство сопоставления фенотипов отдельных особей. Так как фенотипы отражают наследственные свойства особей, то результат этого контроля, выражающийся в естественном отборе наиболее «приспособленных» особей, оказывается довольно сложным. Вследствие указанной связи контролирующий механизм биогеоценоза является косвенным средством контроля: 1) наследственных свойств каждой особи (постольку, поскольку они обнаруживаются в ее фенотипе); 2) качества наследственной информации (поскольку нарушения ее передачи выявляются на фенотипах и 3) качества преобразования наследственной информации в индивидуальном развитии (которое непосредственно определяет свойства фенотипа).
Е. ПРЕОБРАЗОВАНИЕ ИНФОРМАЦИИ В ПОПУЛЯЦИИ
Контроль фенотипов является наиболее существенной предпосылкой для осуществления преобразования информации. Хотя контроль фенотипов имеет основное значение в механизме преобразования информации, мы все же рассматривали его отдельно, так как он проявляется по-разному и, главное, осуществляется в другой системе. Контроль осуществляется внутри всей системы биогеоценоза (на основании сопоставления особей популяции с наличными жизненными средствами), а преобразование, т. е. естественный отбор, протекает внутри самой популяции (на основе сравнительной оценки разных вариантов в этом сопоставлении)
Преобразование обратной информации состоит в изменении состава популяции и вместе с тем сопровождается преобразованием наследственной информации. Результаты преобразования выражаются вполне ясно и четко в форме естественного отбора, который легко доступен как для полевого, так и для экспериментального исследования. Поэтому мы ограничиваем здесь рассмотрение механизма преобразования только действием естественного отбора. Не будем только забывать, что сам естественный отбор протекает под руководящим влиянием взаимоотношений в биогеоценозе, т. е. на основании оценок в разнообразных формах контроля.
Как контроль, так и связанный с ним естественный отбор осуществляются в течение всей жизни особи. Хотя браковка, т. е. элиминация, или отрицательная оценка, возможна лишь однократно, в одном испытании, она может выпасть на любую стадию развития, на любой возраст. Наоборот, переживание и оставление потомства означает положительную оценку во всех многообразных испытаниях на протяжении всей жизни особи. Поэтому элиминация, т. е. отдельная «неудача» при одном лишь испытании имеет в значительной мере случайный характер. С другой стороны, отбор, т. е. положительная оценка при многократных испытаниях в течение всей жизни, получает значение гораздо более достоверного, правильного, почти безошибочного результата. Во всех испытаниях основное значение имеет уровень общей жизненной активности, которая всегда дает особи известные преимущества. Однако формы этой активности могут быть различными и могут получить разную оценку в разных условиях существования.
В этих разных формах жизненной активности и проявляется «приспособленность» организма к данным условиям внешней среды. Способность обеспечить себя жизненными средствами является важнейшим выражением приспособленности. Поскольку в течение жизни любого организма меняются потребности в жизненных средствах и, в частности, как в количестве, так и в качестве пищевых материалов, то и приспособления особи на протяжении всего жизненного цикла также меняются. Все это обеспечивается непрерывным контролем и действием естественного отбора. (Жизненная устойчивость организма определяется, конечно, не только его способностью обеспечить свою жизнь необходимыми для этого средствами. Она выражается и в формах защиты от неблагоприятных внешних влияний. Это — другая сторона приспособленности организмов и мы ее рассмотрим в дальнейшем.)
Естественный отбор принимает различные формы в различных условиях существования и это зависит как от состояния биогеоценоза, так и от особенностей и жизненных потребностей популяции. При изобилии жизненных средств преимущества могут получить и не столь «приспособленные», но зато активные и плодовитые особи. Наоборот, при недостатке необходимейших материалов селекционные преимущества будут на стороне наиболее точно «приспособленных» и даже узко специализированных особей с наиболее экономным обменом, хотя бы и за счет некоторого снижения общего уровня их активности.
Если общее состояние биогеоценоза выражается в среднем известной уравновешенностью и его элементы вполне приспособлены к условиям существования в биогеоценозе, то естественный отбор будет в основном удерживать особей каждой популяции на «нормальном» уровне их приспособленности и тем самым поддерживать состояние равновесия (стабилизирующая форма отбора). Если же в состоянии биогеоценоза происходят сдвиги в опреде-леннном направлении (это может быть обусловлено как климатическими изменениями, так и изменениями в составе его населения, например, вторжением новых форм или вытеснением каких-либо прежних компонентов), то естественный отбор будет идти в пользу известных уклонений, более отвечающих изменившимся условиям существования. Это ведет к новым приспособлениям и перестройке организации (движущая форма естественного отбора).
В масштабе геологического времени такие сдвиги происходят всегда и неуклонно. Однако в любой данный момент почти всякий биогеоценоз можно рассматривать как находящийся в состоянии равновесия. Поэтому стабилизирующая форма естественного отбора отражает условия данного момента, а движущая форма отбора является основой исторического преобразования организмов. Так как история всегда определяется соотношением сил в каждом данном промежутке времени, то и стабилизирующая форма естественного отбора играет свою и притом немаловажную роль в процессе эволюции.
Какие бы формы ни принимал естественный отбор, он всегда приводит не только к изменению состава популяции, но и к перестройке наследственной основы составляющих ее особей, т. е. к преобразованию наследственной информации, и в этом основное его значение. Естественный отбор, т. е. переживание наиболее приспособленных, является положительной оценкой фенотипа. Последний, однако, отражает свойства генотипа (в известных условиях развития особи). Таким образом, вместе с фенотипами отбираются и соответствующие генотипы. В результате получается преимущественное размножение одних фено-генотипов и вытеснение других. Таким образом, меняется состав популяции. При половом размножении, а это следует рассматривать как правило, дело обстоит еще сложнее. При делениях созревания (мейозисе) происходит конъюгация и перераспределение хромосом, а отчасти и обмен их частями в соответствующих парах (перекрест). Таким образом половые клетки получают наследственную информацию от зрелой особи в половинном объеме и в разных вариантах. При оплодотворении происходит новое комбинирование и в результате зиготы получают новую наследственную информацию. Эта последняя является комбинированной информацией от двух особей, апробированных системой контроля биогеоценоза и отобранных из состава данной популяции (табл. 5). Таким образом, хотя естественный отбор протекает по оценке фенотипов, он приводит к преобразованию наследственной информации, т. е. генотипов, В этом случае мы имеем, следовательно, преобразование обратной информации, передаваемой фенотипами на уровне организации особи, в наследственную информацию, передаваемую на молекулярном уровне организации хромосом клетки. Преобразование наследственной информации при половом размножении лишь начинается естественным отбором. Оно продолжается ее перераспределением при созревании половых клеток и завершается новым комбинированием при образовании зиготы. В этом и выражается все преобразование (табл. 6). Последнее протекает внутри самой популяции и захватывает так или иначе каждую особь. Все это заставляет нас отделить факторы преобразования информации, протекающие внутри популяции, от факторов контроля, осуществляющихся внутри всего биогеоценоза в целом, хотя эти последние факторы и имеют решающее значение, как материальная основа всего преобразования.
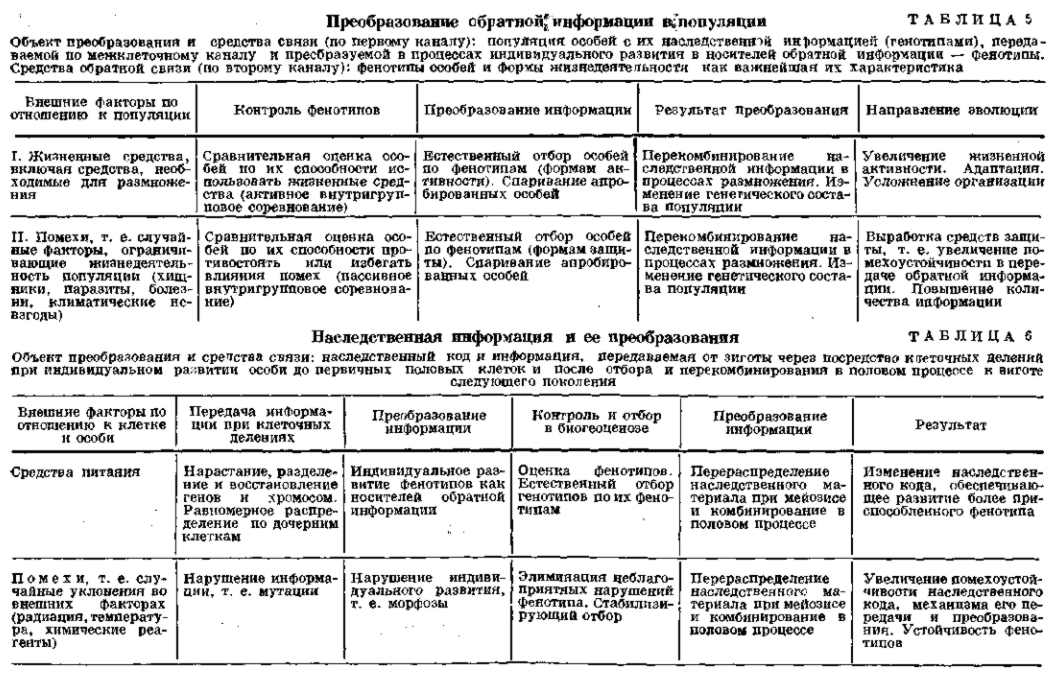
Ж. ЗНАЧЕНИЕ ПОМЕХ В ПЕРЕДАЧЕ ИНФОРМАЦИИ И БОРЬБА С ПОМЕХАМИ
Передача информации часто искажается по самым различным причинам (в звуковой передаче искажения воспринимаются как шумы различного характера). Причины эти могут лежать в построении самой передающей системы, т. е. в несовершенстве использованных средств передачи, но могут вноситься в систему извне. Обычно результаты влияния помех имеют случайный характер — отдельные искажения не могут быть предсказаны и отнесены к определенному моменту времени. Однако после их изучения они могут быть охарактеризованы и оценены в своем среднем значении.
В рассматриваемых нами биологических системах наблюдаются такие же случайные нарушения передачи информации как в первом межклеточном канале передачи наследственной информации, так и во втором канале передачи обратной информации от популяции к биогеоценозу.
1. Помехи в передаче наследственной информации
В передаче наследственной информации возможны нарушения, которые в дальнейшем передаются с такой же точностью, как и унаследованные свойства организма. Эти нарушения, несомненно, связаны с локальными изменениями в строении хромосом. Их источниками являются случайные внешние влияния, особенно радиационные и химические. С повышением температуры частота их возникновения увеличивается согласно правилу Вант-Гоффа (Muller, 1928; Timofeeff-Ressovsky, 1935a). Результаты таких влияний, т. е. искажения передачи наследственной информации, проявляются в виде дискретных наследственных изменений, имеющих также случайный характер. Такие изменения называют мутациями. Случайное появление новых мутаций, конечно, не означает, что явление мутабильности не закономерно. Однако эта закономерность явно статистическая. Случайность мутаций не означает и их недетерминированности. Каждая мутация определяется действием некоторых внешних и внутрисистемных факторов. Однако ввиду сложности всей внутриклеточной системы эти факторы пока еще не расшифрованы. Впрочем, биохимический анализ хромосом и новейшие попытки экспериментального вмешательства в их строение путем химических воздействий приведут, очевидно, в ближайшее время к полному вскрытию механизма возникновения мутаций.
Все такие влияния, искажающие правильность передачи наследственной информации, независимо от их происхождения мы будем называть помехами. Основным их источником мы будем считать внешние воздействия. Эти воздействия могут привести к возникновению мутаций, т. е. к изменению отдельных генов, к их перераспределению через разрыв и перестановку частей внутри хромосом или между хромосомами, к выпадению части в одной хромосоме и удвоению такой же части в ее партнере. Они могут привести и к увеличению числа отдельных или всех хромосом. Во всех этих случаях изменения так или иначе проявляются в процессах преобразования наследственной информации в индивидуальном развитии и в фенотипе данной особи. Наиболее ярко бывают выражены в фенотипах именно генные мутации. Все мутации мы рассматриваем как искажение информации в результате помех.
Большинство мутаций в большей или меньшей мере нарушает жизненные функции организма и при явном их выражении (особенно в гомозиготе) элиминируется в результате отрицательной оценки в контролирующем механизме биогеоценоза. Нередко они, однако, используются в процессе эволюции, особенно в различных комбинациях при исторических изменениях в структуре биогеоценоза. Таким образом мутации (главным образом малые мутации) приобретают иногда положительное значение в определенных условиях существования. Здесь следует отметить, что период созревания половых клеток является и чувствительным периодом по отношению к случайным внешним воздействиям. Таким образом, му-табильность как процесс включается в самые истоки индивидуального цикла жизни и начало первого канала связи. Это обеспечивает контроль наследственных изменений на всем протяжении жизненного цикла особи ( табл. 7).
Очень многие мутации нарушают жизненные функции и являются вредными (особенно летали и мутации с резким физиологи ческим и морфологическим выражением). Такие мутации все же довольно редки.
ТАБЛИЦА 7. Использование результатов помех как материала для эволюции. Возникновение мутаций и их перекомбинирование при образовании половых клеток и оплодотворении. Контроль фенотипов в течение всей жизни особи (начиная с зиготы) в условиях максимальной защиты механизма передачи наследственной информации и ее преобразования от влияния помех

Ясно, что должна быть какая-то защита механизма передачи наследственной информации от ее искажений по меньшей мере в области наиболее существенных ее частей (определяющих жизненно важные структуры и функции).
Теория информации и связи знает следующие основные средства борьбы с помехами. Первое, наиболее простое средство повышения надежности информации заключается в повторении сообщения. Второе — установление связей между элементами сообщения. Третье — увеличение помехоустойчивости через усовершенствование передающей системы. Четвертое — изоляция от помех, т. е. защита передающей системы от проникновения внешних влияний. В передаче наследственной информации использованы все эти средства повышения помехоустойчивости. Наиболее надежным средством повышения помехоустойчивости является установление системных связей в передаваемой информации (частотная модуляция в радиосвязи). Это средство используется в человеческой речи, как в устной передаче (модуляция губами, языком и ротовой полостью на фоне звуковых колебаний голосовых связок), так и в письменной. Фонетические символы — буквы — не комбинируются свободно, а связываются в слова. Посредством логической связи слова соединяются в предложения и т. д.
Чередующиеся пары оснований в молекуле ДНК также не комбинируются свободно. При гибридологическом анализе простейшей единицей наследственной информации оказывается ген. Число качественно различающихся генов исчисляется сотнями и тысячами. Гены представляют собой сложные единицы с глубоко индивидуальной химической структурой. Наличие этих более сложных единиц ограничивает свободу перекомбинирования элементарных химических звеньев (с парами оснований) в молекуле ДНК. Они оказываются связанными в относительно большие и устойчивые комплексы — гены. Таким образом, число возможных комбинаций и, следовательно, количество информации оказывается меньше, чем можно было бы предполагать на основании строения молекулы ДНК. Простейшей единицей наследственной информации является только ген. Максимальное количество наследственной информации определяется поэтому числом генов. В действительности возможное количество информации еще много меньше. Дело в том, что в перекомбинировании генов также нет свободы. Стабильность наследственного механизма, т. е. «помехоустойчивость», увеличивается с помощью дальнейшего усложнения системных связей (между элементами информации). В хромосомах имеются и более крупные, тесно связанные комплексы — сложные гены, супергены и даже целые участки хромосом, передающиеся как непрерывное целое. Во всяком случае, степень сцепления в разных частях хромосом бывает различной (об этом говорит расхождение между цитологической и гибридологической картами хромосом) и это указывает на дифференцировку дальнейших внутрихромосомных систем связей. Во всех этих случаях связываются, очевидно, элементы наследственной информации, имеющие особо важное значение в развитии всего организма как целого.
Еще более общее значение имеет соединение всех этих наследственных элементов и комплексов в целостных хромосомах. Каждая хромосома представляет с точки зрения теории информации сложное сообщение со многими «логическими» связями. Наконец, механизм митотического клеточного деления обеспечивает совместную передачу полного набора хромосом. Следовательно, и все хромосомы одного набора оказываются связанными меж собой через этот механизм. Таким образом составляется полная информация об индивидуальных качествах данного генотипа. Внутрихромосомные связи, т. е. строение генов и явления сцепления, играют, таким образом, большую роль в обеспечении надежности передачи наследственной информации от зиготы ко всем клеткам организма вплоть до первичных половых клеток.
Однако надежность этой передачи не исключает возможности ее изменения и использования таких изменений в эволюции. Такие изменения очень редки при обычной митотической передаче от клетки к клетке. Они, однако, относительно легко осуществляются в процессах созревания половых клеток (в мейозисе). Конъюгация и перекрест хромосом могут вести к разрыву сцепления и обмену гомологичными частями парных хромосом. Могут быть и нарушения, ведущие к хромосомным перестройкам. Наконец, не исключены и изменения в структуре генов. Таким образом, стабильность наследственного механизма не исключает возможности его изменения, особенно во время чувствительного периода образования половых клеток.
Самым простым средством повышения надежности информации является (применяемое и в технике связи) ее повторение. В передаче наследственной информации использовано и это средство повышения помехоустойчивости. Прежде всего под этим углом зрения можно рассматривать повторение генов с одинаковым или сходным выражением. Комбинирование таких «полигенов» приводит к количественным градациям в известных признаках организации и в этом, быть может, их основное значение, как материала для эволюции (Mather, 1941). С другой стороны, однако, самый факт множественного повторения генов в полигенных системах должен приводить к известной устойчивости этих систем как средств передачи наследственной информации. Отдельное нарушение (т. е. изменение одного гена) не отзывается сколько-нибудь заметно на функции полигенной системы в целом, а следовательно, и не вносит особых изменений в фенотип. Это — очень выгодное для эволюции сочетание некоторой лабильности и возможности градуальных изменений с довольно высокой устойчивостью. При перестройках хромосом возникают иногда повтор-ности частей, называемые дупликациями. В гетерозиготной форме дупликации содержат трехкратное, в гомозиготной форме — четырехкратное повторение некоторого участка хромосом. Так как в нормальных хромосомах слюнных желез дрозофилы встречаются повторения участков со сходным строением (Bridges, 1935), то весьма вероятно, что дупликации нередко приобретали положительное значение в процессе эволюции. И в этом случае их значение могло лежать в том же счастливом сочетании известной лабильности (возможности отдельных мутаций) с высокой устойчивостью наследственной передачи вследствие наличия повторных участков. Наследственная информация в целом не нарушается при наличии отдельных «ошибок» в одном из повторных сообщений. Все это, однако, лишь частные случаи повторности некоторых, быть может особенно существенных, элементов наследственной информации. Зато всеобщее значение имеют повторения одинаковых хромосом в диплоидном организме. Значение диплоидности как защиты от повреждающего действия мутаций общеизвестно — в гетерозиготном состоянии большинство мутаций не имеет заметного выражения. Диплоидность поэтому, с одной стороны, допускает наличие мутационных изменений и, с другой стороны, подавляет их фенотипическое выражение. В этом опять ясно выражается положительная роль повтор-ностей в передаче наследственной информации. С одной стороны, возможность ее нарушений (что может быть использовано в процессе эволюционных преобразований) и, с другой стороны, все же достаточно надежная ее передача.
Еще выше должна быть помехоустойчивость полиплоидного организма. В этом случае многократное повторение тех же хромосом, несомненно, сильно увеличивает надежность передачи наследственной информации и ее преобразования, а следовательно, и фенотипическую стабильность организации. Многократно отмечавшаяся стабильность полиплоидов растений в более суровых климатических условиях является, очевидно, выражением этой «помехоустойчивости». Вместе с тем полиплоидия обеспечивает большую потенциальную пластичность организма в эволюционных процессах.
Несомненно, что в повышении стабильности механизма наследственной передачи большую роль сыграли и последние два средства увеличения помехоустойчивости — усовершенствование передающей системы и изоляция, т. е. защита от внешних влияний.
Стабильность передающей системы поддерживается химической устойчивостью молекул ДНК, их связью в генах и хромосомах, весьма совершенным механизмом клеточных делений. Кроме того, весь наследственный материал («код») хорошо изолирован ядерной оболочкой и телом клетки от каких бы то ни было внешних влияний. Все такие влияния опосредствуются прежде всего организмом в целом с его многочисленными регуля-торными механизмами («гомеостатом») и затем самой клеткой, представляющей собой также элементарный регуляторный механизм. Таким образом, снимается вся специфика внешних воздействий.
Помехоустойчивость наследственной информации оказывается поэтому исключительно высокой. В ее обеспечении сыграли свою роль все возможные средства: повторность сообщений (в хромосомах и в парах хромосом), связанность знаков и их комплексов в целых сообщениях (в генах, хромосомах и их наборах), стабильность всего механизма передачи (химическая устойчивость и механизм клеточного деления) и изоляция от помех (через посредство регуляторных механизмов организма и клеток). К этому следует добавить, что и процессы преобразования информации в индивидуальном развитии также хорошо защищены от «ошибок» вследствие высокой селективности восприятия (применяемой и в технике) в частях развивающейся особи, специфичностью реакций преобразования (как и в технике), а также сложной системой внутренних связей (гомеостатом).
2. Помехи в передаче обратной информации через фенотип.
Если передача обратной информации осуществляется в различных формах активного воздействия особей данной популяции на биогеоценоз, то все нарушения активности, т. е. жизнедеятельности отдельных особей, мы, очевидно, должны обозначить как результат «помех». Источниками помех могут быть и в этом случае внутрисистемные причины — дефекты морфофизиологичес-кой организации фенотипов. Однако основными «помехами» являются неблагоприятные влияния факторов неорганической среды (климата и т. п.) ив особенности воздействия биотических факторов — болезней и паразитарных инвазий, которые могут сильно снизить активность пораженных особей, а могут и прервать их жизнь, и хищников, которые также являются препятствием для свободного проявления жизнедеятельности и вместе с тем истребляют более или менее значительное число особей. Все эти внешние воздействия на популяцию имеют такой же случайный характер, как и любые типичные «помехи». Естественно, что и против этих помех вырабатываются свои средства защиты.
Помехоустойчивость в передаче обратной информации достигается также различными средствами. Большую роль и здесь играет повторение сообщения. Хотя каждая особь является целостным носителем информации, это не значит, что последняя неразложима. Фенотип особи составляется из очень многих признаков, более или менее тесно спаянных. Связность сообщения здесь очень велика и фенотип весьма устойчив, в особенности в наиболее существенных чертах своей организации. И в этой организации особи встречаются повторности, такие как антимерия и метамерия. В каждой ткани — многократные повторности клеточных элементов. Все это вместе взятое должно способствовать устойчивости особи и надежности обратной информации от популяции в целом.
Каждая особь является самостоятельным и дискретным носителем информации. Хотя каждая особь глубоко индивидуальна, т. е. отличается от любой другой особи по многим признакам и даже по всей организации в целом, однако в своих существенных чертах каждая особь повторяет любую другую особь данной популяции и даже данного вида. Таким образом, допуская варианты информации, их основа остается в общем одной и той же. Именно в своей наиболее существенной части информация повторяется и в каждой особи. Поэтому информация, исходящая от всей популяции в целом, оказывается в своей сущности вполне застрахованной даже при гибели многих особей.
Своеобразным проявлением повторности особей данного вида в наиболее существенных чертах своей организации являются также различные формы полиморфизма. В некоторых случаях они могут послужить основой для развития следующей ступени повторности географических и экологических форм и, наконец, привести к новым видовым повторностям. Явления полиморфизма требуют довольно сложного генетического механизма, регулирующего закономерный характер повторности. Гораздо проще это достигается при частичном (по полам, по сезонам или по циклам размножения) или полном отказе от спаривания при половом процессе. Партеногенез, апогамия, вегетативное размножение приводят к групповой однородности, т. е. к повторению одних и тех же фенотипов (в линиях, клонах и т. п.). Все эти явления, по-видимому, связаны с наличием значительных «помех», именно — с большой истребляемостью (различными факторами как биотическими, так и абиотическими). Однако во всех этих случаях повышение качества информации осуществляется за счет значительного уменьшения ее количества (определяемого многообразием форм). Несомненно, что эти явления не служат показателем прогрессивной эволюции, а связаны скорее с застоем на известном уровне организации.
О значении связей между элементами информации, т. е. признаками в пределах особи, мы уже говорили. Однако возможны и зависимости между особями. В некоторых случаях наблюдается, хотя бы периодически, совместная жизнедеятельность особей разного пола попарно или стадами. Иногда особи объединяются для совместной жизни или по меньшей мере для выведения потомства в более или менее значительные колонии. Очень часто животные вообще держатся, перемещаются и кормятся стадами, стаями или косяками. Во всех этих случаях совместное кормление и размножение способствуют, очевидно, более успешному и надежному проявлению жизненной активности (т. е. передаче обратной информации), более успешной защите от хищников.
Жизненная устойчивость особей имеет, конечно, первостепенное значение для безошибочной передачи информации о состоянии популяции. В этом выражается совершенство самой системы передачи. Жизненная устойчивость достигается в основном морфофизиологической слаженностью всей организации и наличием сложной системы регуляторных механизмов. Еще большее значение имеет создание некоторого избытка активности. Избыток активности проявляется у высших позвоночных в особенности в виде известного запаса в организации центральной нервной системы, который играет исключительную роль в прогрессивной эволюции высших млекопитающих (и в происхождении человека). В этих случаях помехоустойчивость достигается не только уклонением от опасностей, осторожностью в поведении, но и активной борьбой со всеми вредными влияниями.
Наконец, большое значение в передаче обратной информации имеет также изоляция от помех. Особи являются единственными свободными носителями информации. Поэтому они непосредственно сталкиваются с самыми разнообразными внешними воздействиями. Защита от таких влияний приобретает первостепенное значение. В защите от абиотических влияний, которые могли бы нарушить нормальную жизнедеятельность особей, основным является их общая устойчивость. Однако большое значение имеют и методы пассивной защиты, т. е. изоляции. Сюда относятся временный покой растений и животных, что у последних связано нередко с использованием разного рода укрытий (анабиоз, спячка). Животные пользуются и временными укрытиями для защиты от холода или от летнего зноя. Растения изолируются от влияния засухи сокращением испарения. От хищников и вредителей вырабатываются, кроме средств активной борьбы (у животных), также меры пассивной защиты. Таковыми являются твердость покровов, шипы и колючки (как у растений, так и у животных), иногда и ядовитые вещества тканей (у растений) или выделений желез (у животных). У животных к этому прибавляются — маскировка (защитные окраска, форма), предупреждающие сигналы защищенных форм и мимикрия, или маскировка под защищенные формы (ложная информация). Наконец, у высших животных основную роль играют инстинкты, позволяющие им избегать встречи с хищниками, рано их распознавать и скрываться от них. Во всех этих случаях организм защищается от хищников или вредителей путем изоляции от их воздействия. Таким образом популяция в значительной мере охраняется от чрезмерного влияния помех в передаче информации. В эволюции высших животных известную роль в этом смысле играли прогрессивное развитие защитных средств кожных покровов — чешуя, волосы, но кроме того и общее увеличение размеров тела.
Важным фактором изоляции у растений является одеревенение тканей и развитие твердой коры. Прогрессивная эволюция растений всегда подавлялась деятельностью травоядных животных и суровыми условиями континентальных климатов. Развитие «помехоустойчивости» у прогрессивных видов многократно приводило к возникновению древесных форм, защищенных зимним покоем от мороза и иногда также летним покоем от засухи. Древесные формы, как правило, хорошо защищены и от высших травоядных животных. Однако древесные растения вследствие медленной смены генераций всегда отставали в своей эволюции от травянистых и вытеснялись затем новыми древесными растениями, развившимися из опередивших их травянистых. Так было с древовидными хвощами и папоротниками, а также с хвойными и саговыми, на смену которым пришли древесные покрытосеменные.
Из сказанного, конечно, не следует, что эволюция древесных форм определялась в основном влиянием повреждающих факторов. Основное значение и здесь, как и в эволюции травянистых форм, имела борьба за максимальное использование световой энергии для фотосинтеза.
Мы определили помехи как случайные внешние влияния. Если, однако, они являются постоянными элементами биоценоза, то их действие проявляется вполне закономерно. Они входят в состав контролирующего механизма и, следовательно, через вну-тригрупповое сопоставление особей влияют на ход естественного отбора. Таким образом, они участвуют в преобразовании информации и определяют также направления эволюционного процесса. Эти направления сводятся, однако, в данном случае не столько к общему прогрессу организации, сколько к частным адапта-циям, т. е. к выработке средств защиты от помех. Впрочем, достигнутая мера помехоустойчивости сама определяет затем и возможности прогрессивной эволюции.
3. ЗНАЧЕНИЕ НЕПОСРЕДСТВЕННОГО ВЛИЯНИЯ ВНЕШНИХ ФАКТОРОВ В ПРОЦЕССЕ ЭВОЛЮЦИИ
Как бы организмы ни были защищены от помех в передаче информации, все же эта защита далеко не абсолютная. Поэтому влияние случайных внешних факторов всегда сказывается на фенотипах популяции, а также и на процессе эволюции. Нарушения в передаче наследственной информации, выражающиеся в фенотипе как мутации, всегда проходят через механизм контроля в биогеоценозе и в большинстве случаев элиминируются. Иногда они, однако, приобретают положительное значение (главным образом в комбинациях), преобразуются и используются в прогрессивной эволюции. Это возможно лишь при безотказной работе механизма контроля. Так как мутации представляют нарушения нормальной организации и нормальных функций организма, то бесконтрольное накопление мутаций ведет всегда к дезинтеграции и распаду организации — деградации. Если контроль снимается лишь в отношении некоторых органов или признаков, то деградируют — редуцируются — именно эти органы или признаки. Если контроль снимается в отношении большинства органов (в упрощенной среде), это ведет к почти всеобщей дегенерации (у паразитов). Если контроль снимается в отношении признаков, потерявших значение при переходе в совершенно иную обстановку, то распадаются именно эти признаки, например, средства защиты, окраска и другие признаки при одомашнении. Таким образом, непосредственное влияние нарушений в передаче наследственной информации, т. е. помех как случайных факторов, всегда имеет дезорганизующий характер.
Помехи в передаче обратной информации, т. е. случайные факторы, нарушающие, сокращающие или прерывающие жизнедеятельность особей известной популяции, сами по себе также оказывают дезорганизующее влияние на передачу информации и на структуру всей популяции. Информация сокращается количественно и может быть сильно искажена. В случае избирательного уничтожения по полам или возрастам (например, преимущественное истребление самок, молоди или яиц) это может оказать резко отрицательное влияние на состав популяции. Чем более мощным является уничтожающий фактор, тем более общий характер приобретает элиминация особей. Сокращение численности компенсируется тогда только увеличением плодовитости. Во всяком случае никакие помехи как случайные внешние воздействия сами по себе не являются факторами прогрессивной эволюции. Входя, однако, как постоянный элемент в состав биогеоценоза, они становятся вместе с тем частью его контролирующего механизма и через процесс естественного отбора принимают участие в эволюционных преобразованиях по пути развития частных адаптации (преимущественно средств защиты). Таким образом, воздействия внешних факторов на механизм передачи информации, несомненно, имеют известное значение и являются источником изменений, лежащих в основе эволюционного процесса. Однако их использование возможно только через посредство контролирующего и регулирующего механизма биогеоценоза.
Непосредственные воздействия внешних факторов на преобразующий механизм индивидуального развития могут повлечь за собой и изменения фенотипа в тех случаях, когда они выражают некоторые уклонения от нормы. Наоборот, обычные влияния внешних факторов входят в условия нормального течения процессов индивидуального развития. К ним организм «приспособлен», и они участвуют в самой реализации этой приспособленности.
При нормальном преобразовании наследственной информации в индивидуальном развитии происходит, несомненно, сложная цепь биохимических и морфологических изменений. Эти изменения происходят во взаимодействии частей развивающегося организма и во взаимодействии всего организма в целом с факторами внешней среды. При этом, как правило, сам наследственный материал (код) сохраняется, или, точнее, воспроизводится в неизменном виде, и притом не только в зачатковом пути и в первичных половых клетках, но и в соматических. Это видно по закономерному воспроизведению унаследованных признаков не только при половом размножении, но и при вегетативном. В последнем случае источником воспроизводимого материала, за счет которого строится дочерняя особь, являются соматические клетки, которые уже претерпели преобразование (почкообразование у мшанок, оболочников, различные формы вегетативного размножения у растений), И все же из дифференцированного соматического материала развиваются нормальные полноценные особи., которые даже с особой точностью воспроизводят наследственные свойства родителей.
Непосредственно внешние факторы затрагивают лишь механизм преобразования информации в индивидуальном развитии. Их влияние не может быть передано прямо на наследственный материал, который достаточно устойчив и защищен от повреждающих факторов благодаря наличию регуляторных механизмов клетки и организма в целом. Косвенное влияние через процессы нарушенного клеточного обмена не может быть специфическим, оно может выразиться лишь в возникновении «неопределенных» случайных мутаций. Если такое влияние скажется на половой клетке или зиготе, то это и будет мутация, которая может приобрести положительное значение лишь в известных условиях и во всяком случае должна пройти через обычный механизм контроля. Если же это влияние обнаруживается на более поздней стадии развития и коснется лишь части организма, то это будет соматическая мутация, которая может быть использована (через механизм контроля) лишь в редких случаях при вегетативном размножении или у организмов с поздним обособлением зачаткового материала (губки, гидроиды, растения).
Как уже указывалось, «неопределенность» мутаций есть результат сложного опосредствования всех внешних влияний, при котором вся специфика последних снимается. Это вовсе не значит, что мутирование лишено какого бы то ни было направления. Наоборот, мы знаем, что некоторые мутации возникают гораздо чаще других. Несомненно, что это определяется разной устойчивостью отдельных генов, а также систем передачи и отдельных звеньев механизма преобразования наследственной информации. Это может вызвать и картину направленного мутирования. Если некоторая мутация связана с изменением в морфогенной системе, выходящим за пределы нормального реагирования, то дальнейшие изменения в том же направлении должны происходить с гораздо большей легкостью. Если пороговый уровень преодолен, защитный механизм преобразования прорван, то открывается свободный канал для дальнейших изменений. Конечно, это может привести к эволюционным преобразованиям лишь в том случае, если это направление мутирования получит благоприятную оценку в контрольном механизме биогеоценоза. Вероятно, многие мутации являются специфическим ответом на некоторые внутриклеточные продукты обмена; возможно, что они имеют даже до известной степени «приспособительный» характер. Однако такие реакции могут рассматриваться только как внутрисистемные «приспособления». Они могут лишь «случайно» оказаться благоприятными для всего организма в целом. Их судьба решается во всяком случае только в процессах контроля и естественного отбора.
Мы должны здесь особо отметить, что в мутировании нет свободы изменений каждого гена и ни в какой мере нет равновероятности мутаций. Мутабильность определяется строением всей передающей системы и ограничивается наличием сложной системы связей и взаимозависимости не только в хромосомах, но и во всей клетке в целом. Кроме того, фенотипическое выражение мутации определяется и ограничивается наличным механизмом индивидуального преобразования.
Наследственный код очень хорошо защищен от обычных внешних влияний. Это имеет исключительно большое значение для точной передачи наследственной информации из поколения в поколение и для возможности контролируемого ее преобразования в процессе эволюции. Однако это имеет не меньшее значение и для возможности контроля и регуляции процессов индивидуального развития. Этот контроль и регуляция осуществляются, несомненно, при самом активном участии наследственного материала. Возможность такой регуляции сама уже предполагает известную стабильность контролирующего и регулирующего механизмов. Развивающийся организм может таким образом контролировать и изменения, индуцируемые внешними факторами.
Необычные уклонения во внешних факторах могут нарушать преобразование информации в течение процессов онтогенеза и могут привести к необычным изменениям фенотипа. Фенотип оказывается искаженным (морфозы). Он не отражает свойств генотипа в обычных условиях его реализации. Поэтому и обратная информация оказывается ложной. Это крайне затрудняет контроль в биогеоценозе. Обычно такие уклонения (морфозы) элиминируются по фенотипам (хотя их генотипы могут быть вполне полноценными) .
Внешние влияния могут, следовательно, в некоторых случаях нарушить механизм преобразования информации в индивидуальном развитии. Так как это преобразование происходит только в соме, то его нарушение не касается зачатковых клеток (а если оно и коснется их, то это будет мутация). Фенотип оказывается измененным и, как правило, нарушенным в своих нормальных (для всей популяции) соотношениях с факторами среды. Обычно это ведет через оценку в контрольном механизме биогеоценоза к элиминации. Однако в случае, если данное уклонение получило даже положительную оценку и особь приступила к размножению, ее потомство оказывается «нормальным», неизменным, так как наследственная информация (т. е. ее код) не была нарушена. Только в одном случае — если влияние среды обусловлено индивидуальной восприимчивостью к данному влиянию, т. е. наследственными свойствами особи, новая форма реагирования может при ее положительной оценке в контрольном механизме биогеоценоза быть использована в эволюции и войти в состав «нормы» реакций данного вида организмов.
Таким образом, наследование признаков, приобретенных при жизни особи, фактически невозможно, так как это «приобретение» касается только преобразования информации в данной особи и гибнет вместе с этой особью. Наследственный материал не был затронут этим преобразованием и остался неизменным.
Конечно, можно себе представить такое нарушение развития и в особенности такое изменение обмена, которое окажется вновь в роли «помехи», искажающей передачу наследственной информации, т. е. нарушающей структуру наследственного кода. Это выразится опять-таки в форме мутации. Нет, конечно, никаких оснований ожидать, что фенотипическое выражение этой новой мутации окажется сходным с выражением фенотипа, возникшего в результате нарушения в преобразовании информации, т. е. в процессах индивидуального развития. Так как и факторы воздействия различны, и реагирующий материал (части развивающегося организма, с одной стороны, и наследственный материал хромосом — с другой) совершенно различен, то сходство в фенотипах измененной особи и ее потомства было бы исключительно невероятным событием. Как правило, результаты непосредственного действия внешних факторов на фенотип развивающейся особи не могут быть использованы в процессе эволюции. Измененная особь, даже пройдя через контроль и естественный отбор по фенотипам, не сможет передать своих свойств потомству, так как ее генотип остался неизменным или изменился в совершенно ином направлении.
Внешние факторы могут влиять на формообразование особи и могут его изменять, однако они не могут его контролировать, так как каналы обратной связи от организма к внешней среде не могут быть использованы. Во внешней среде нет механизма для преобразования информации и передачи управляющих сигналов на развивающуюся особь (в системе популяции механизм преобразования имеется, но он эффективен только за пределами особи).
Изменение родителей в течение их онтогенеза, т. е. изменение, вновь приобретенное под влиянием внешних факторов и не детерминированное изменением наследственного материала (т. е. мутацией), может повлечь за собой какое-то изменение потомства (через плазму или в процессах созревания половых клеток). Однако реализация этого изменения находится вне какого-либо контроля со стороны родителей, их фенотипа, или их наследственного материала, так как нет обратной связи от потомства к родителям.
Обратная информация от детей к родителям, контроль и регуляция со стороны родителей устанавливаются только у высших позвоночных — птиц и млекопитающих. Они выражаются в явлениях воспитания, т. е. индивидуального формирования основ поведения. Конечно, это воспитание не ведет к каким-либо наследственным изменениям.
Таким образом, все приобретаемые изменения могут сыграть какую-то роль в эволюции только при условии, что они не только индуцируются внешними факторами, но одновременно имеют в своей основе хоть какие-либо изменения унаследованной нормы реакции. В этом случае измененный фенотип отражает также и изменение генотипа. Только при этом условии оценка фенотипа в контролирующем механизме эволюции адекватна оценке его генотипа. Поэтому и естественный отбор по фенотипам приведет к преобразованию генотипа, т. е. наследственной информации, вливающейся в популяцию как новое ее приобретение.
Определенное, направленное изменение в процессе эволюции не может быть результатом прямого воздействия внешних факторов. Оно всегда основывается на случайных наследственных изменениях (мутациях) и определяется сложной системой контроля и преобразования (естественным отбором) при тесном взаимодействии внутренних и внешних факторов данного биогеоценоза. Так как мутации в своей основе случайны, то тем большее значение имеет косвенное влияние внешних факторов через механизм контроля фенотипов в биогеоценозе. Результатом этого влияния и является преобразование наследственной информации в процессе естественного отбора и комбинирование апробированных генотипов при половом размножении.
Для нас важно подчеркнуть, что возможность успешного контроля и регуляции эволюционного процесса определяется безупречным функционированием всего регулирующего аппарата. Прежде всего это касается безошибочной передачи наследственной информации на всем протяжении жизни особи, т. е. от зиготы до первичных половых клеток (свободным для элементарных преобразований остается лишь небольшой период созревания половых клеток. Второе условие — надежное преобразование этой информации в индивидуальном развитии. Фенотип должен быть точным выразителем генотипа (для данных условий развития и жизни). Только в этом случае контроль и естественный отбор по фенотипам означают контроль и отбор генотипов и могут обеспечить их адекватное преобразование в биогеоценозе. Поэтому защита индивидуального развития фенотипа от случайных внешних влияний и ненаследуемость соответствующих изменений являются необходимыми условиями для возможности регулируемого преобразования организма на пути его последовательного приспособления к обычным факторам внешней среды (биогеоценоза) и на пути закономерной его эволюции вообще. Приспособление организма к внешней среде возможно только через посредство контроля, отбора и преобразования случайных изменений, возникающих при созревании половых клеток. Говоря о значении внешних факторов, нельзя не упомянуть и о значении приспособительного реагирования. Мне многократно приходилось подчеркивать большую положительную роль непосредственного приспособления организмов к изменениям во внешней среде (1939а,б, 1942 и др.). Такое индивидуальное приспособление входит в развиваемое здесь представление о преобразовании наследственной информации в индивидуальном развитии. Оно всегда основано на унаследованных нормах реакций. Способность к приспособительным реакциям возникает на основе мутационных изменений нормы реакции и создается в результате естественного отбора наиболее благоприятных форм реагирования. Развиваясь таким образом, способность к приспособительным реакциям приобретает огромное значение как база для возникновения новых дифференцировок. Частные реакции, выделившиеся на основе общей способности приспособительного реагирования, могут получить значение постоянного приобретения (в данных условиях). В результате действия стабилизирующей формы естественного отбора такие реакции всегда входят в установившуюся «норму» и приобретают устойчивую наследственную основу. Это не требует больших наследственных преобразований, так как каналы для развития данной модификации уже выработались в предшествующей истории (при отборе форм реагирования). Необходимо лишь возникновение мутаций, которые привели бы к замене данного внешнего раздражителя внутренним. При большой специфичности унаследованных реакций и крайне малой специфичности большинства морфогенети-ческих влияний такая замена совершается с большой легкостью путем подстановки сопутствующих влияний и изменения порогового уровня реактивности формообразовательного материала.
Это — незначительные количественные изменения, которые всегда обеспечиваются обычной мутабильностью наследственного кода.
И. ЕСТЕСТВЕННЫЙ ОТБОР И ЕГО ФОРМЫ
Естественный отбор осуществляется на основании сравнительной оценки особей в контролирующем механизме биогеоценоза. Поэтому действие естественного отбора, а следовательно, и общий характер преобразования наследственной информации определяются состоянием самого биогеоценоза.
Обычно экологи рассматривают соотношения в биогеоценозе как состояние подвижного равновесия. И это правильно, так как это действительно характеризует данный исторический момент, в котором производится исследование эколога. Однако, с другой стороны, мы точно знаем, на основании палеонтологических, а частью и прямых исторических данных, что состав биогеоценозов неуклонно менялся, а вместе с тем менялись и характеризующие их виды организмов. Мы должны учитывать оба аспекта: с одной стороны, состояние относительного равновесия в данном дифференциале времени и факторы, поддерживающие это равновесие, и, с другой стороны, смещение точки равновесия в определенном направлении и факторы, вызывающие соответственные изменения организмов.
I. Сдвиги в соотношениях между популяцией и внешней средой нарушают точную приспособленность «нормальных» особей. Они ведут к естественному отбору некоторых уклонений от «нормы» (мутаций при агамном размножении и сложных их комбинаций при половом размножении), которые в измененных условиях существования получили какие-то преимущества в жизненном состязании с представителями прежней «нормы». Это — классическая дарвиновская форма естественного отбора, ведущая через суммирование и интеграцию уклонений к изменению существующих и развитию новых приспособлений. Новые приспособления возникают обычно в результате дифференцировки уже существующих, на основе принципов разделения и смены функций. Таким образом, происходит непрерывное приспособление организма к меняющимся условиям существования и постепенное усложнение его строения. Я назвал эту форму естественного отбора движущей формой отбора. Преобразующее действие естественного отбора основывается при этом на использовании таких нарушений («ошибок») в передаче наследственной информации, которые после преобразования в индивидуальном развитии (т. е. в фенотипах) получают положительную оценку в контролирующем механизме биогеоценоза (в новых его условиях). Апробированные уклонения от нормы скрещиваются, комбинируются и размножаются, давая все новую наследственную информацию и, после преобразования в онтогенезе, новые фенотипы, вливающиеся в популяцию. В каждой генерации эти фенотипы проходят контроль и через естественный отбор и скрещивание преобразуют генетическую структуру и фенотипическое выражение особей всей популяции.
В результате этого изменяется унаследованная норма и ее код, изменяются процессы преобразования в индивидуальном развитии и изменяются фенотипы как носители обратной информации.
II. В условиях равновесия между популяцией и внешней средой (в биогеоценозе), т. е. в условиях данного исторического момента, естественный отбор поддерживает существование и размножение установившейся «нормальной» (для данного момента) организации. Отбор идет на основании селекционного преимущества нормальных особей перед всеми уклонениями от этой приспособленной нормы. Эта форма естественного отбора ведет к сокращению изменчивости, т. е. к «нормализации» популяции, ее особей и признаков, и к выработке более устойчивых механизмов наследования и индивидуального развития особей. Я назвал эту форму естественного отбора стабилизирующим отбором.
Преобразующее действие стабилизирующего отбора основывается при этом на использовании небольших уклонений (мутаций) в наследственной информации, которые, не сказываясь заметно на фенотипе сформированной особи, приводят к более надежному воспроизведению установившейся нормы. Это означает увеличение устойчивости наследственного аппарата и механизма преобразования наследственной информации против различных случайных влияний, которые часто нарушают нормальное течение индивидуального развития. Обратная информация о масштабе таких нарушений передается в биогеоценоз, где и происходит оценка нормальной жизнедеятельности особей в установившихся условиях существования. Преобразование этой информации через естественный отбор завершается размножением апробированных «нормальных» особей, обладающих наиболее устойчивым механизмом наследственности и индивидуального развития.
Результатом действия стабилизирующего отбора является увеличение «помехоустойчивости», т. е. надежности передачи и преобразования наследственной информации. Это достигается всеми возможными средствами борьбы с помехами — повторностью генов (полигены) и их комплексов (дупликация, гетеро- и полиплоидия), установлением систем связей (увеличение сцепления в супергенах, в участках хромосом и в целых хромосомах), а также усовершенствованием и изоляцией как механизма наследственной передачи, так и механизма ее преобразования в индивидуальном развитии. Вследствие элиминации всех случайных уклонений в фенотипах, в установившихся условиях существования, стабилизирующий отбор ведет к максимальной защите наследственной информации от случайных внешних влияний. Защита механизма преобразования информации достигается опосредствованием этих влияний в развивающемся фенотипе, именно регуляторным характером как внутриклеточных взаимозависимостей, так и всего формообразовательного аппарата. Стабилизирующий отбор ведет, таким образом, к созданию механизма авторегуляции и к возможной в данном случае автономизации процессов индивидуального развития.
Вместе с тем это ведет, конечно, и к максимальной устойчивости результатов индивидуального развития, т. е. сформированных фенотипов зрелых особей, а следовательно, и к максимальной надежности обратной информации, носителями которой являются эти фенотипы. Количество информации уменьшается с увеличением однородности («нормализацией») популяции, однако ее качество значительно повышается. Увеличивается внутренняя связь (система корреляций) между элементами организации, и каждая особь приобретает максимальную целостность. Каждая особь оказывается полноценным носителем почти всей информации, характеризующей данную популяцию. Повторность информации захватывает почти всю организацию. При максимальной устойчивости механизма преобразования информации в автономизированном индивидуальном развитии фенотип дает наиболее точное выражение свойств генотипа. Поэтому и контроль в биогеоценозе, осуществляющийся путем сопоставления фенотипов, и механизм преобразования (естественный отбор фенотипов и перекомбинирование соответствующих им генотипов) наследственной информации в биогеоценозе должны работать с наибольшей надежностью (т. е. «безошибочно»).
Как видно, стабилизирующая форма естественного отбора ведет к совершенствованию всего регулирующего механизма эволюции.
Движущая форма естественного отбора меняет приспособления организма по мере изменений в условиях его существования, перестраивает и усложняет его функции и строение. Стабилизирующая форма отбора в каждый данный момент закрепляет достигнутые результаты, связывает их в целостную систему и обеспечивает максимальную надежность их воспроизведения
К. НАПРАВЛЕНИЕ ЭВОЛЮЦИОННОГО ПРОЦЕССА В ЦЕЛОМ
В целом эволюция органического мира идет по пути приспособления к данным условиям существования и по мере смены и усложнения последних ко все далее идущей дифференциации функций и усложнению организации (алломорфоз). К существенным чертам эволюции следует отнести также общее повышение организации и уровня жизнедеятельности организмов, которые отмечают узловые моменты больших эволюционных преобразований (ароморфозы А. Н. Северцова), ведущих к адаптивной радиации данных форм на новом уровне. Не преуменьшая значения таких больших качественных преобразований, мы должны отметить, что известное повышение уровня жизнедеятельности и активности организма входит как элемент генеральной линии эволюции и в повседневные явления прогрессивной адаптации живых существ к условиям их существования. Мы говорим именно о генеральной линии эволюции, оставляя в данном случае в стороне нередкие уклонения в направлении дегенерации (катаморфоз) и недоразвития (гипоморфоз), которые могут привести к известной деспециализации и вновь дать начало прогрессивной эволюции в новом направлении. К генеральной линии эволюции следует, однако, отнести и общее увеличение устойчивости организма на всех стадиях его развития — устойчивости половых клеток и зиготы с их аппаратом передачи наследственных свойств организма, устойчивости механизма индивидуального развития и устойчивости фенотипа организма во всех его возрастах. Эта устойчивость является результатом действия стабилизирующей формы естественного отбора. Однако естественный отбор и, в частности, его стабилизирующая форма регулирует не только историческое преобразование организмов и их наследственнной основы. В результате его направляющего действия происходит также закономерная перестройка самого механизма эволюции.
Эта эволюция эволюционного процесса строится на той же материальной основе, как и вся эволюция, т. е. на мутациях, которые проходят тот же индивидуальный контроль в биогеоценозе (на основе сравнительной, т. е. внутригрупповой оценки). Здесь проявляется значение как движущей, так и в особенности стабилизирующей формы отбора. Однако особое значение имеет в дальнейшем межгрупповое соревнование популяций в темпах и качествах своего преобразования. В этом соревновании происходит вытеснение одних популяций другими, что можно назвать и групповым (популяционным) отбором. Во всех случаях исторического изменения самого механизма эволюции начало этих преобразований лежит всегда в популяциях, однако их оценка переносится затем на межгрупповое соревнование в темпах и качестве эволюционных преобразований между популяциями, экологическими и географическими подразделениями вида, между видами, родами и целыми филогенетическими ветвями, как отряды, классы и даже типы. Вытесняя друг друга с командных позиций, они показывают, следовательно, не только преимущества своей организации, но и преимущества того механизма эволюции, с помощью которого им удалось обогнать другие формы в темпах своего приспособления, в повышении организации и общего уровня активности и завоевать таким образом руководящее положение среди живых существ данной эпохи. Перестройка механизма эволюции выражается в следующем:
1. Под руководящим влиянием естественного отбора и межгруппового состязания вырабатываются оптимальные формы наследственной и ненаследственной изменчивости, регулируется уровень мутабильности по разным признакам и вырабатывается гибкий механизм перекомбинирования мутационных изменений (в хромосомах, между хромосомами и в парах хромосом).
2. В процессе эволюции действием естественного отбора в межгрупповом состязании меняются средства распространения наследственных изменений внутри популяции и вида в целом через организацию мейозиса, использование полового процесса (в разных формах) и частичное ограничение нанмиксии при разных формах изоляции.
3. В процессе эволюции, под влиянием стабилизирующей формы отбора, увеличивается помехоустойчивость и совершенствуются каналы передачи наследственной информации, т. е. усложняется строение наследственного кода введением повторности и развитием системы связей в хромосомах, увеличивается надежность равномерной передачи наследственных структур при клеточных делениях.
4. В процессе эволюции, под влиянием стабилизирующей формы естественного отбора, вырабатывается механизм преобразования наследственной информации (во взаимодействии с внешними факторами) в индивидуальном развитии особи. Увеличивается устойчивость этого механизма через выработку регуляторных систем, которые играют большую роль в прогрессивной автономизации развития и в дальнейшей прогрессивной эволюции.
5. В процессе эволюции увеличиваются активность организмов и специфика их жизнедеятельности, что обеспечивает более надежную передачу обратной информации и ускорение темпов эволюции.
6. Под влиянием стабилизирующей формы естественного отбора усложняется система внутренних связей (корреляций) во всей организации и увеличивается устойчивость особей (фенотипов) как носителей обратной информации.
7. В процессе эволюции меняются формы контроля (борьбы за существование) и естественного отбора, т. е. механизма преобразования информации в биогеоценозе. Вырабатываются наиболее эффективные комбинации внутригруппового и межгруппового соревнования и форм отбора в условиях панмиксии и ее ограничений.
8. В межгрупповом состязании происходит естественный отбор наиболее совершенных механизмов эволюции, обеспечивающих наиболее быстрые ее темпы.
Регулирующий механизм эволюции неуклонно усложняется в процессе самой эволюции, совершенствуются средства передачи информации в обоих направлениях и механизмы ее преобразования, но основные принципы функционирования системы контроля и преобразования (на двух ее уровнях — в онтогенезе особи и в наследственном изменении популяции) остаются в общем теми же самыми.
На основании всего сказанного о регуляции и контроле в эволюционном процессе, мы приходим к следующим выводам:
1. Эволюция может рассматриваться как автоматически регулируемый процесс. Объектом управляемых изменений является, несомненно, популяция как первичная эволюирующая единица. В качестве регулирующего механизма выступает, очевидно, биогеоценоз, включающий данную популяцию. Связь между биогеоценозом и популяцией осуществляется в двух направлениях и на двух разных уровнях организации. Прямая информация о преобразованиях в биогеоценозе передается через посредство апробированных особей путем их размножения. Каждая дочерняя зигота получает наследственную информацию от своих родителей и передает ее путем клеточных делений вплоть до первичных половых клеток зрелой особи. Наследственная информация преобразуется в процессах индивидуального развития. Результатом преобразования является сформированный фенотип, отражающий (соответственно условиям среды) свойства генотипа на уровне организации особи. Обратная информация передается от популяции к биогеоценозу через посредство фенотипов. Средством связи является здесь жизнедеятельность каждой особи, влияющая на соотношение сил в биогеоценозе. Передача информации осуществляется и в этом случае с помощью символов, однако эти символы — признаки фенотипа — не имеют ничего общего с символами наследственного кода (генами).
Фенотипы особи данной популяции проходят в течение всего своего жизненного цикла непрерывную цепь контрольных испытаний в биогеоценозе. Эти испытания могут иметь лишь два исхода — гибель на какой-либо стадии развития и жизни (элиминация) или переживание вплоть до оставления потомства (естественный отбор). Элиминация имеет всегда случайный характер. Естественный отбор является закономерным выражением результатов сложной системы контроля в биогеоценозе. Отбор протекает на основе сравнительной оценки фенотипов внутри популяции (внутригрупповое соревнование особей) и является основным механизмом преобразования обратной информации в биогеоценозе. Это преобразование начинается на уровне организации особи и приводит к перестройке наследственной информации в системе клетки.
Поскольку фенотипы правильно отражают свойства генотипа и являются одновременно также носителями наследственной информации (в первичных половых клетках), вместе с отбором по фенотипам происходит и отбор генотипов. Это вносит изменения в генетический состав популяции. Половой процесс вносит еще дальнейшие изменения — при созревании половых клеток происходит случайное перекомбинирование и распределение наследственного материала. При оплодотворении происходит объединение наследственной информации, полученной от двух апробированных особей в одной зиготе. Последняя является, таким образом, носителем преобразованной наследственной информации. Основой этого преобразования является естественный отбор.
2. Обычные факторы внешней среды входят как необходимый элемент в преобразующие механизмы прямой информации (в индивидуальном развитии) и в преобразующий механизм обратной информации (контроль в биогеоценозе). Однако необычные, случайные воздействия могут оказать неблагоприятное влияние на наследственную информацию, на механизм преобразования этой информации в развитии особи, на проявления жизненных функций, т. е. передачу обратной информации, и на преобразующий механизм биогеоценоза. Все такие внешние влияния случайного характера можно объединить и рассматривать в качестве помех, нарушающих правильность работы регулирующих механизмов эволюции. Элиминация таких случайных нарушений развития и жизнедеятельности особей приводит к естественному отбору' (стабилизирующему) наиболее помехоустойчивых систем передачи информации и ее преобразования.
Нарушения в передаче наследственной информации выражаются в мутациях. Элиминация вредных мутаций приводит к развитию более надежного механизма этой передачи. Повторность генов (полигены) и их комплексов (дупликация), особенно диплоидия и полиплоидия, являются средствами повышения надежности информации. Дальнейшими средствами увеличения помехоустойчивости являются внутренние связи в системе передачи — строение генов из многих химических единиц, связь генов в супергенах и их сцепление в целых хромосомах и, наконец, объединение целого набора хромосом в механизме клеточного деления. Помехоустойчивость поддерживается также локализацией наследственного кода внутри тела клетки под защитой регуляторных механизмов клетки и всего организма в целом.
Помехоустойчивость механизма преобразования информации |в индивидуальном развитии поддерживается селективностью и специфичностью форм реагирования, организацией систем внутренних связей (корреляций), регуляторными механизмами формообразовательных систем и физиологическими регуляциями всего организма в целом (гомеостатом). Надежность обратной информации обеспечивается наибольшей слаженностью организации, системой коррелятивных взаимозависимостей, определяющих максимальную устойчивость фенотипов, а также повторностью всех существенных элементов организации в каждой особи. Кроме этого, существенную роль играют и средства изоляции, т. е. пассивной защиты от всех неблагоприятных воздействий внешних факторов.
3. В прогрессивной эволюции организма по пути адаптации и усложнения организации основную роль играет движущая форма естественного отбора. Одновременно с усложнением фенотипа как носителя обратной информации происходит при этом и усложнение наследственного аппарата как механизма передачи прямой информации и основы его преобразования в индивидуальном раз витии.
В прогрессивной эволюции организма как целого, в развитии его устойчивости, ведущую роль играет, однако, стабилизирующая форма отбора. Через элиминацию всех случайных нарушений фенотипа она ведет к организации наиболее надежных каналов связи, к наибольшей помехоустойчивости как в передаче наследственной информации, так и в ее преобразовании в индивидуальном развитии и в реализации фенотипов как носителей обратной информации.
Движущая форма отбора меняет строение организма по мере изменений в его условиях существования, она перестраивает систему его адаптации, усложняет его функции и поднимает всю организацию на высший уровень жизнедеятельности.
Стабилизирующая форма отбора в каждый данный момент закрепляет достигнутые результаты, связывает их в устойчивую целостную систему и обеспечивает максимальную надежность ее воспроизведения.
4. В процессе прогрессивной эволюции, на основе системы контроля и регуляции в биогеоценозе, через внутригрупповое состязание и естественный отбор происходит не только направленное, закономерное изменение организмов, генеральная линия которого определяется повышением уровня их активности, но и закономерная перестройка и совершенствование самого механизма эволюции. В этой перестройке играет известную роль как движущая, так и в особенности стабилизирующая форма естественного отбора, ведущая к усовершенствованию механизмов передачи информации. Как надстройка над внутригрупповым состязанием особей, в этом случае приобретает, однако, исключительное значение и межгрупповое состязание в темпах и качестве эволюционных преобразований. Это состязание ведет к вытеснению одних групп другими, что можно назвать и групповым отбором.
11. ЗНАЧЕНИЕ СТАБИЛИЗИРУЮЩЕГО ОТБОРА В ПРАКТИКЕ РАСТЕНИЕВОДСТВА И ЖИВОТНОВОДСТВА
Не подлежит никакому сомнению, что естественный отбор играет немалую и притом еще далеко не вполне оцененную роль в развитии наших культурных растений и домашних животных. Хорошо, если направление этого отбора соответствует требованиям человека и используется им (отбор на морозоустойчивость, на засухоустойчивость или устойчивость против грибных и бактериальных заболеваний и т. п.) как фактор, содействующий селекции наиболее производительных сортов и пород. В большинстве случаев, однако, естественный отбор наиболее жизнеспособных особей не вполне совместим с искусственным отбором более продуктивных линий. В этом случае приходится создавать особые условия воспитания и содержания для повышения жизненной устойчивости получаемых сортов и пород. Кроме того, человек идет по пути создания наиболее подходящих условий для максимального выявления ценимых им свойств, т. е. для возможного повышения продуктивности. В обоих случаях это означает сознательное модифицирование форм. Так как, однако, противодействие естественного отбора непрерывно ведет к изменению созданных человеком форм (главным образом в сторону исходной), то во избежание «вырождения» приходится вести борьбу за чистоту сорта или породы путем жесткой браковки уклонений от установленного «нормального» стандарта. Элиминация уклонений должна вести к известной стабилизации сорта или породы.
Во многих случаях уровень агротехники явится фактором, который сам по себе, без сознательного участия человека, приведет через стабилизирующий отбор к изменению наследственных свойств организма. В условиях высокой агротехники, если урожайность зерна повысится до максимальной, генотипически более урожайные особи дадут максимальное выявление своих свойств. Доля этих особей в продукции урож;ая будет выше, чем относительное число самих особей. Если 10% всех особей дадут лишь на 10% большее число семян, то в общем урожае будет уже почти 11% зерен более урожайных особей. Так же будет возрастать доля более урожайных особей и в следующих поколениях. Эволюция в условиях высокой агротехники будет идти всегда в направлении наследственного повышения урожайности, т. е. в направлении стабилизации достигнутого уровня. /
В условиях низкой агротехники (недостаток удобрения, влаги; густой посев) генетически наиболее урожайные особи не смогут выявить полностью своих свойств. Наоборот, они могут оказаться в худшем положении, чем менее урожайные особи, так как требуют для своего нормального развития более значительного питания. Генетически наиболее мощные особи окажутся в условиях наибольшего минерального и водного голодания. Их урожайность упадет ниже, чем у других особей, или они окажутся даже совсем бесплодными (см. известные исследования В. Н. Сукачева о сравнительной плодовитости разных линий одуванчика в редких и густых посадках, 1927, а также о сравнительной мощности развития и плодоношения при свободном стоянии и при конкуренции у Fesiuса sulcata, 1935). То же самое касается не только продукции семян, но и клубнеобразования, отложения сахара и других резервов в корнях и т. п., так как особи наиболее продуктивные окажутся в этих условиях наиболее истощенными, дадут меньше семян, и их удельный вес в популяции будет неуклонно падать.
Стабилизирующий отбор по низкоурожайному фенотипу (модификации) приведет через элиминацию более урожайных линий к быстрому снижению урожайности, т. е. к наследственному вырождению сорта. В условиях активной конкуренции произойдет не только общее снижение урожайности (как модификация), но будет идти и естественный отбор на дальнейшее ее снижение (см. И. Шмальгаузен, 1939а, стр. 186, 197 и далее). Произойдет наследственная стабилизация низкой урожайности, возникшей как модификация в условиях низкой агротехники.
Все эти факты более или менее известны, в них нет ничего принципиально нового. Искусственный стабилизирующий отбор, в смысле широкой браковки всех уклонений от установленного стандарта, давно применялся, и его эффективность является лишним доводом в пользу того, что и в естественных условиях стабилизирующий отбор, идущий, конечно, с гораздо большей жесткостью, должен иметь большое значение.
Можно ли, однако, дать, на основании всего сказанного, руководящие указания практике для более успешной и быстрой стабилизации новых сортов и пород? Мне кажется, что некоторые выводы напрашиваются сами собой.
Ясно, что практикуемый в данных случаях инцухт в пределах одной линии совершенно непригоден, так как процесс гомозиготизации генотипов катастрофически сокращает основной материал для стабилизирующего отбора.
Необходимо не только постоянное скрещивание для поддержки возможного разнообразия генотипов, но и применение всяких других методов для повышения мутабильности форм (например, резкие изменения внешних условий) даже тогда, когда желательные качества нового сорта уже получены.
В особенности совершенно необходимо заботиться о возможной гетерогенности материала в тех случаях, когда желательные свойства нового сорта имеют хотя бы частично модификационный характер. Стабилизация модификаций возможна лишь при наличии максимально гетерогенного исходного материала. Только в этом случае жесткая браковка всех уклонений от установленного стандарта может помочь достигнуть наследственной устойчивости новых форм. Понятно, эта браковка должна производиться в конечном счете в тех условиях культуры, в которых данный сорт предполагается использовать.
Экспериментальное сопоставление результатов браковки уклонений от частично модифицированного и вновь установленного стандарта на основе гетерогенного материала, с одной стороны, и чистого сорта — с другой, помогло бы нам кстати и разрешить вопрос о механизме наследственной стабилизации новых сортов. С точки зрения ламаркистов, допускающих непосредственное фиксирование ненаследственных изменений в ряде поколений, стабилизация модификационных изменений должна идти в пределах чистого сорта с неменьшей скоростью, чем в пределах гетерогенного сорта. Между тем стабилизирующий отбор должен во втором случае дать гораздо более быстрый результат, чем в опытах с чистыми сортами.