
Проблемы Эволюции |
Глава (С.50-62) из книги: А.Ю.Розанов. Закономерности морфологической эволюции археоциат и вопросы ярусного расчленения нижнего кембрия. М.: Наука, 1973. 164 с.
Глава (С.50-62) из книги: А.Ю.Розанов. Закономерности морфологической эволюции археоциат и вопросы ярусного расчленения нижнего кембрия. М.: Наука, 1973. 164 с.
В 1920 г. Н. И. Вавилов впервые сформулировал в кратком виде закон гомологических рядов. Установленный на основе громадного фактического материала, собранного как самим Н. И. Вавиловым, так и многочисленными его учениками и коллегами, закон, как ни странно, не был в то время воспринят большинством специалистов-зоологов, ни тем более палеонтологов. Хотя, как сейчас становится ясно, открытие закона гомологических рядов стоит в ряду крупнейших открытий нашего столетия.
Лишь Д. Н. Соболев, В. А. Догель, Л. С. Берг, Бульман (Bulmann) и ряд других исследователей смогли оценить значение сделанного открытия, так как в той или иной степени развивали сходные представления в своих работах.
Н. И. Вавилов рассматривал закономерности гомологической изменчивости на современном материале, т. е. на одном хронологическом срезе. Нетрудно представить себе, что, несмотря на неполноту палеонтологической летописи и специфику сохранности палеонтологического материала, детальное изучение ископаемых организмов в этом аспекте может дать несомненный эффект благодаря возможности учета фактора времени.
Многие палеонтологи, занимавшиеся систематикой, интуитивно использовали в решении проблем классификации закономерности гомологической изменчивости групп.
История развития представлений о системе археоциат является удивительным в этом отношении примером. Начиная с первых систематик (Taylor, 1910), большинство последующих попыток систематизации так или иначе невольно делалось на основе анализа закономерностей гомологической изменчивости (Simon, 1939, Краснопеева, 1955; Вологдин, 1956 и др.; Журавлева, 1960; Debrenne, 1964 и др.). Для примера здесь приводятся таблицы П. С. Краснопеевой и Симона (табл. 1, 2).
Таблица 1. Периодическая система родов и подродов (по Simon, 1939)


В настоящее время исследования гомологической изменчивости у ряда ископаемых групп уже показали несомненную ценность такого подхода к разработке их систематик (Поярков, 1969; Липина, 1971 и т. д.). Попытки исследования гомологической изменчивости у археоциат были предприняты мною, начиная с 1962 г. (Розанов, 1964; Розанов, Миссаржевский, 1966; Розанов, 1969, 1971а), и дали возможность, оценив некоторые закономерности в эволюции археоциат, произвести уточнения в их систематике. В последнее время аналогичные работы по Monocyathida проводила О. Г. Окунева (рис. 74).

Рис. 74. Гомологические ряды отряда Monocyathida (по О.Г.Окуневой)
Исследования гомологической изменчивости ископаемых организмов позволяют, прежде всего, оценить таксономическое значение признаков и отличить яркие, но несущественные в систематическом плане признаки, от менее ярко морфологически выраженных, но гораздо более существенных.
Археоциаты — вымершая еще в кембрии группа организмов, поэтому функциональное значение отдельных элементов скелета не совсем ясно. Мы можем лишь рассуждать по аналогии с близкими группами: простейшими и губками. Вместе с тем удивительное морфологическое тождество различных элементов скелета у разных групп археоциат и однозначная их топография дает возможность, как нам кажется, говорить о гомологичности этих элементов скелета и гомологической изменчивости вообще.
Отметим, что в данном случае речь идет о гомологии на макроуровне (на уровне морфологических признаков). По-видимому, нет необходимости, анализируя гомологическую изменчивость любой группы (ископаемой или ныне живущей), смешивать различные уровни организации и пытаться разобрать вопросы гомологии на морфологическом уровне с точки зрения гомологии на уровне клеточной или внутриклеточной организации. Колоссальные сложности, стоящие на пути увязки различных уровней организации — от гена до организма в целом, естественно проявляются в оценке гомологии. Нет сомнений, что работы по последней проблеме совершенно необходимы, однако вспомним, что для целей изучения закономерностей гомологической изменчивости и решения конкретных задач растениеводства Н. И. Вавилову потребовался огромный материал, но изученный на одном уровне.

щелкните, чтобы увеличить
Рис. 75. Таблица гомологических рядов археоциат (по Розанову, 1966)
1 — Archaeolynthus; 2 — Tumuliolynthus; 3 — Rabdocyathella; 4 — Capsulocyathus; 5 — Fransuasaecyathus; 6 — Fransuasaecyathidae gen. nov.; 7 — Uralocyathella; 8 — Dokidocyathus; 9 — Incurvocyathus; 10 — Alphacyathus; 11—Dokidocyathella; 12 — Kaltatocyathus; 13 — Papillocyathus; 14 — So-anicyathus; 15 — Zhuravlevaecyathus; 16 — Kidrjasocyathus; 17 — Tchojacyathus; 18 — AjacicyathuS; 19 — Orbicyathus; 20 — Archaeocyathellus; 21 — Urcyathus; 22 — Archaeofungia; 23 — Leptosocyathus 24 — Leptosocyathus; 25 — Gordonicyathus; 26 — Compositocyathus; 27 — Gordonifungia; 28 — Ethmophyllum; 29 — Ethmophyllum; 30 — Afiacyathus; 31 — Tumulocyathus; 32 — Tumulocyathellus; 33 —Porocyathus; 34 — Kiiacyathus; 35 — Annulofungia; 36 — Vologdinocyathus; 37 — Russocyathella; 38 — Ladaecyathus; 39 — Erbocyathus; 40 — Urcyathella; 41 — Pretiosoeyathus; 42 — Tegerocyathus; 43 — Nochoroicyathus; 44 — gen. nov.; 45 — Pectenocyathus; 46 — Thalamocyathus; 47 — Formosocyathus; 48 — Kotuytcyathus; 49 — Jakutocyathus; SO — Lenocyathus; 51 — Geocyathus; 52 — Korde-cyathus; 53 — Coscinocyathus; 54 — Asterocyathus; 55 — Salairocyathus; 56 — Coscinocyathellus; 57 — Tumulocoscinus; 58 — Alataucytahus; 59 — Asterotumulus; 60 — Porocoscinus; 61 — Tomocyathus kundatus, 62 — Tomocyathus operosus; 63 — Kazyricyathus; 64 — Mrassucyathus
В этом смысле Н. И. Вавилов (1935, стр. 47) совершенно справедливо писал: «Конечно, не во всех случаях, а тем менее у разных родов изменение выявляется всегда одними и теми же, т. е. гомологическими генами. Одинаковые изменения фенотипического порядка могут быть вызваны и разными генами. Поскольку систематика имеет дело с признаками, поскольку при обозрении растительного и животного мира мы имеем дело пока в основном не с генами, о которых мы знаем очень мало, а с признаками в условиях определенной среды, более правильно говорить о гомологических признаках».
Касаясь примеров гомологических рядов у археоциат на разных таксономических уровнях, оговоримся, что понятия вид, род и т. д. у археоциат (как, по-видимому, во многих случаях в палеонтологии) носят достаточно условный характер и отражают наше представление о соподчинении таксонов, реальный ранг которых нами не может быть строго доказан.
В настоящее время понятие рода у археоциат более или менее однообразно у всех авторов. Начиная именно с этой категории, таксономические различия имеют качественную характеристику (присутствие — отсутствие признаков), в то время как при понимании (разграничении) видов и более мелких категорий употребляются наряду с некоторыми качественными, преимущественно количественные отличия.
Исследований по изменчивости видов археоциат проводилось очень мало, поэтому примеры гомологических рядов на внутривидовом уровне в настоящее время привести практически невозможно. Эта работа только начинается. Здесь лишь можно отметить, что, например, виды Coscinocyathus dianthus Born, и С. rojkovi Vol. имеют кубки как конической, цилиндрической, так и тарельчатой формы. При этом нет сомнений, что данный признак не может рассматриваться как признак видового различия (т. е. мы не рассматриваем эти виды сборными), так как экземпляры с тарельчатыми и коническими кубками встречаются в одном слое, с той лишь разницей, что конические формы приурочены чаще к небольшим биогермным телам, а тарельчатые — к межбиогермным, более глинистым фациям.
При сравнении набора видов отдельных родов, особенно многочисленных по видовому составу, легко увидеть практически полную повторяемость признаков. Для примеров приведем таблицу признаков видов таких известных и широко распространенных родов, как Aldanocyathus Voron. и Robustocyathus Zhur. (табл. 3).

Повторяемость многих признаков хорошо видна из этой таблицы. Следует добавить, что ряд признаков, в том числе и указанных в таблице могут встречаться в самых различных комбинациях по 2—3—4 и т. д. И в этом случае мы можем фиксировать такие сочетания у видов, как одного, так и другого рода.
Наиболее отчетливо наличие гомологических родов прослеживается на уровне рода, что частично было освещено ранее (Розанов, Миссаржевский, 1966, (см. рис. 45); Розанов, 1969, 1971). В настоящее время в результате получения многочисленных новых материалов (Коршунов, Журавлева, Розанов, 1969; Окунева, 1969; Беляева, 1969; Воронин, 1969; Розанов, 1969; Debrenne, 1969; Дебренн, Розанов, 1972 и т. д.) можно дать более расширенный вариант таблицы гомологических рядов (см. рис. 77).
Сравним для примера два ряда родов семейств Ajacicyathidae и Cyclocyathellidae.
AldanocyathusVoron. Kellericyathus Roz.
Robustocyathus Zhur. Gordonicyathus Zhur. [Thalamocyathus]
Ajacicyathus Bedf. Denacyathus Zhur.
Mongolocyathus Voron. Compositocyathus Zhur.
Sibirecyathus Vol. Gordonifungia Roz.
Эти два семейства отличаются строением внутренней стенки, т. е. у семейства Cyclocyathellidae присутствуют кольцевые образования. При сравнении видно (рис. 76), что рол Aldanocyathus имеет пористые перегородки и несколько рядов пор на внутренней стенке, аналогичным образом у рода Kellericyathus — пористые перегородки, а внутренняя стенка имеет две-три поры на интерсептум плюс кольцевые пластины. У рода Robustocyathus — пористые перегородки и один ряд пор на внутренней стенке, а урода Gordonicyathus снова прибавляются лишь кольцевые пластины (признак семейства). Род Mongolocyathus имеет непористые перегородки и один ряд пор на внутренней стенке, то же имеет и Compositocyathus, но с кольцевыми пластинками. Род Ajacicyathus имеет непористые перегородки и несколько рядов пор на внутренней стенке, то же мы видим и у рода Denacyathus. И, наконец, у рода Sibirecyathus наблюдаются синаптикулы, как и у рода Gordonifungia. Точно такой же ряд можно наблюдать и у этмофиллид. Например, с пористыми перегородками, непористыми перегородками и с синаптикулами (рис. 77).

Рис. 76. Гомологические ряды (сем. Ajacicyathidae и Cyclocyathellidae) 1 — Aldanocyathus; 2 — Robustocyathus; 3 — Ajacicyathus; 4 — Mongolocyathtis; 5 — Sibirecyathus; 6 — Kellericyathus; 7—Qordonicyathus; 8 — Denaecyathus; 9 — Compositocyathus; 10—Gordonifungia
Сравнивая отдельные семейства, принадлежащие даже разным надсемействам, мы легко обнаружим и там те же наборы. Например, в надсемействе Annulocyathaсеа есть семейство Gloriosocyathidae, имеющие в своем составе формы с пористыми перегородками (Gloriosocyathus Roz.), с непористыми перегородками (Rossocyathella Zhur.) и с синаптикулами (Ring-ifungia Korsch.).
Сравнивая же надсемейства по другим признакам, можно видеть, что почти каждое надсемейство имеет формы с простой пористостью внутренней стенки, с козырьками или чешуями на внутренней стенке, и наконец, с разнообразными каналами (этмофиллоидные стенки).
Не менее удивительная картина полной повторяемости структур наружной стенки может быть прослежена при сравнении даже таких крупных категорий, как подотряды. Легко убедиться (рис. 77), что везде имеются формы с простой пористостью наружной стенки, с тумуловыми (причем и простыми и бугорчатыми), с козырьками у устьев пор и со всеми видами дополнительных оболочек. Известные мне случаи приведены в таблице (рис. 77 и табл. 4).
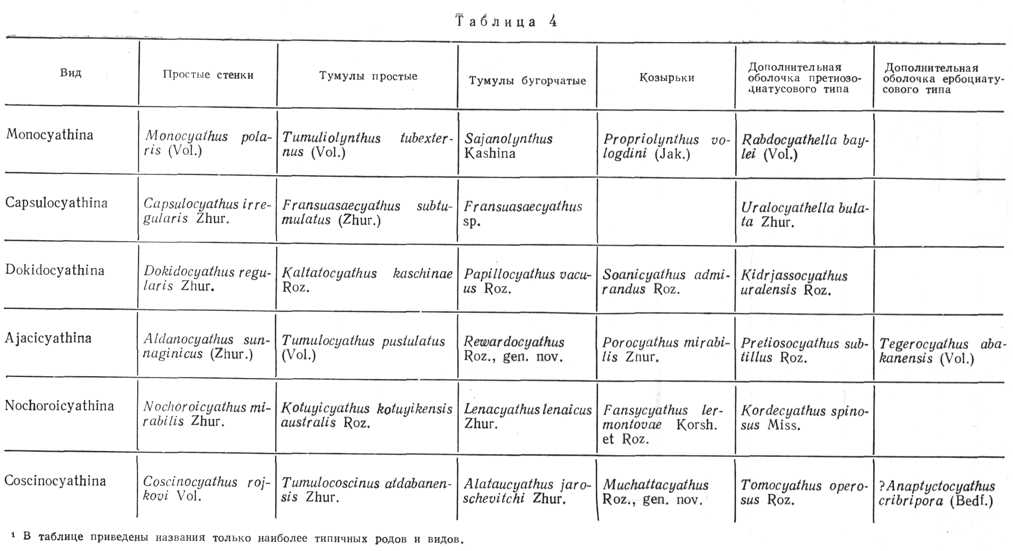
Выявление гомологических рядов у археоциат (как, впрочем, по-видимому, и в любой другой группе) позволяет с большой степенью достоверности предсказать морфологические особенности форм, которые могут быть еще найдены. Именно таким образом были предсказаны, а затем обнаружены Tumulifungia Zhur., Hupecyathellus Roz. и др. (Розанов, Миссаржевский, 1966; Журавлева, 1967). В настоящее время можно с тем же основанием предположить, что будут еще найдены формы, например, имеющие дополнительную оболочку претиозоциатусового или ербоциатусового типа, перегородками с синаптикулами и внутренней стенкой всех известных типов (простой, чешуйчатой, кольцевой, или этмофиллоидной). Иными словами — в таблице гомологических рядов не заполненные ячеи говорят о возможности нахождения форм, морфология которых соответствовала их месту в данной ячейке.
Интересно, что среди известных по литературным данным и находящиеся в коллекциях форм практически нет таких, которые не могли бы быть помещены в предлагаемую таблицу.
Исключение составляют три рода (Cryptaporocyathus Zhur., Ethmolynthus Zhur., Salopicyathus Vol.), о которых уже говорилось ранее в разделе «Морфология». Повторю, что Ethmolynthys, по-видимому, внутренняя стенка двустенного археоцита, Salopicyathus — единственный известный экземпляр, по-видимому, со вторично утолщенной наружной стенкой. Особое место занимает Cryptaporocyathus — реальный очень хорошо изученный род. Здесь может быть ряд объяснений. Во-первых, это представитель Monocyathida, и не исключено, что морфологические возможности моноциатид несколько отличались от Ajacicyathida. Во-вторых, не исключено, что пока мы просто не находили аналогов такой стенки у Ajacicyathida, но что они будут найдены. И в-третьих, может быть Cryptaporocyathus не является археоциатом, а принадлежит какой-то иной близкой группе примитивных Metazoa или еще точнее Archeata (см. Журавлева, Мягкова, 1971).
Это не означает, конечно, что дальнейшая работа по уточнению морфологии археоциат не приведет к увеличению количества ячей. Как раз наоборот, нужно думать, что в дальнейшем мы вынуждены будем строить не только более дробную, но и более объемлющую таблицу. Я исхожу здесь из того, что несколько лет тому назад мы не различали, например, группы с претиозоциатусовой и ербоциатусовой оболочками или с тумуловыми и козырьковыми наружными стенками, и каждая из этих пар рассматривалась как единое целое. Более детальное изучение морфологии скелета археоциат может привести, например, к выделению самостоятельных групп со стенками, которые сейчас условно именуются «молоточки» (Журавлева, 1970) и т. д.
Само по себе существование гомологических рядов дает возможность определения границ возможных морфологических преобразований, которые, естественно, регламентируются конструктивными потенциями данной группы.
Наличие закономерностей гомологической изменчивости заставляет думать, что эволюция археоциат (как, по-видимому, и других групп) может происходить, и очевидно происходит, по принципу исчерпания возможностей морфологических преобразований, в связи с чем выявление путей реализации является нашей следующей задачей.

Щелкните, чтобы увеличить
Рис. 77.
Обратите внимание на карандашные пометки. Это последователи и ученики А.Ю.Розанова после открытия новых видов археоциат заполняли пустые клетки его периодической таблицы (!)