
Проблемы Эволюции |
Из книги: Эволюционная биология. Материалы конференции "Проблема вида и видообразования". Томск: Томский государственный университет, 2001. - Т.1. - С. 128-137.
[из книги: Эволюционная биология. Материалы конференции "Проблема вида и видообразования". Томск: Томский государственный университет, 2001. - Т.1. - С. 128-137]
Эволюционная идея в биологии базируется на осмыслении фундаментальных философских принципов, а именно - значения и соотношения "внутреннего и внешнего" в развитии или, по биологической терминологии, - автогенеза и эктогенеза. Основатель эволюционного учения гениальный Ж.Б. Ламарк впервые ввел эти два принципа в биологическое мировоззрение и воплотил их в двух законах: 1)градации (автогенез) и 2) приспособительной эволюции (эктогенез). Ч. Дарвин и его последователи, включая и современных "неодарвинистов", проповедуют лишь второй принцип (эктогенез), оценивая естественный отбор как главную причину эволюционного развития. Другой крайностью является абсолютизация "внутреннего" в эволюции, или автогенеза. Наиболее яркий приверженец этого подхода - Л.С. Берг.
С позиций современной биологии становится очевидным, что эволюционный процесс на всех этапах осуществляется при непрерывном взаимодействии "внутреннего и внешнего" [1]. Именно это взаимодействие отвечает принципам системного подхода, когда система (внутреннее) функционирует или развивается только под контролем среды (внешнее). В данной статье предполагается оценить проблему "внутреннего", то есть организации видовой системы, и прежде всего её генетической компоненты, на современном уровне развития биологии. Эволюционная судьба вида как генетической системы может быть разной в зависимости от его структурно-функциональной организации. Системный подход предполагает наличие в видовой системе инвариантной (мономорфной) структуры - видового генома и вариантной (полиморфной) компоненты - видового генофонда. При этом вводим постулат, что реорганизация видового генома может привести к развитию системы или видообразованию, тогда как трансформация видового генофонда является основой поддержания функциональной стабильности вида (видового гомеостаза). По определению [2], основой функционирования системы являются гомеостатические (равновесные) отношения. Основой развития являются противоречивые (неравновесные) отношения. Если в процессе функционирования качество системы не меняется, то такой тип её поведения требует сохранения определенной устойчивой структуры.
Видовой генофонд - полный набор вариантных состояний генов (аллелофонд) и хромосомных структур (инверсионный полиморфизм и др.), характерный для популяций в пределах видового ареала. При этом не всегда генетический полиморфизм относится к адаптивному типу, а лишь тогда, когда он обеспечивает стационарность видовой системы в условиях экоклиматических градиентов среды обитания. В настоящее время находит подтверждение точка зрения, оценивающая адаптивный генетический полиморфизм в терминах стационарной видовой системы и популяционно-генетического гомеостаза, исключающих возможность градуальной трансформации видов [З]. Альтернативная позиция большинства современных неодарвинистов [4, 5], оценивающих пространственно-временные частотные флуктуации полиморфных вариантов генов и хромосомных инверсий как проявление постепенной видовой дивергенции или филетической эволюции, представляется нам ошибочной. При этом сторонники синтетической теории эволюции обычно используют следующие аргументы:
1) в пространстве видового ареала происходят постепенные замещения альтернативных аллельных и инверсионных вариантов (клины), что, по их мнению, в конечном счете может привести к полной дизрупции; подобные процессы могут характеризовать и темпоральные частотные сдвиги;
2) реальные частоты аллельных и инверсионных генотипов не соответствуют распределению Харди-Вайнберга (наблюдается значительный дефицит гетерозигот);
3) обнаруживаются ассоциативные взаимодействия между разными инверсиями и аллелями (linkage disequilibrium). Необходимо отметить, что подобная «эволюционная» интерпретация обычно является следствием крайне редукционного подхода, когда отдельные аллели или инверсии, обнаруживающие подобные тенденции, сознательно вырываются из контекста оценки всего кариофонда или аллелофонда.
Напротив, комплексный и тщательный анализ генофонда и кариофонда вида показывает наличие у видов исключительной стационарности адаптивного полиморфизма, отражающей высокую степень популяционно-генетического или, точнее, видового гомеостаза. При этом выявляются популяционные системы, обладающие мощным интегрирующим эффектом на фоне дифференциальной частотной динамики по отдельным аллелям или инверсиям. Было убедительно доказано, что популяционно-генетический гомеостаз реализуется у видов с адаптивным генетическим полиморфизмом через пространственно-временную частотную динамику инверсий и аллелей и динамичность их ассоциативных связей. При этом в генокомплексах (сочетаниях разнохромосомных инверсий и аллелей) всегда существуют генотипические варианты (отдельные гены или хромосомы), играющие роль интеграторов генофонда, обеспечивающих видовое единство в тех случаях, когда некоторые инверсионные варианты или аллели имеют тенденции к дифференциации генофонда за счет ассоциативных связей или дефицита гетерозигот [6].
Несмотря на некоторые модификации взглядов неодарвинистов, касающиеся предварительных условий и скорости видообразования, все они базируются на постулате: при видообразовании реорганизуется накопленный ранее генофонд вида. Градуализм зиждется на частотном преобразовании полиморфной популяции с последующей фиксацией наиболее адаптированных аллельных или хромосомных вариантов. Этот методологически ошибочный, противоречащий системному подходу дарвиновский принцип «разновидность - суть возникающий вид», подхваченный современными неодарвинистами в их обычной трактовке «от полиморфизма к видообразованию», не выдерживает критики. Из теории систем следует, что абсолютное преобладание гомеостатических структур (адаптивного генетического полиморфизма) приводит только к сохранению системы, но не к ее преобразованию (видообразованию).
После фундаментальных работ Ю.П. Алтухова и Ю.Г. Рычкова [3, 7], в которых наряду с конструктивной критикой градуального видообразования были сформулированы принципы преобразования видов, основанные на реорганизации инвариантной (мономорфной) части видового генома, в последние десятилетия XX века были получены оригинальные данные, подтверждающие и развивающие эту концепцию [6, 8, 9, 10, 11].
Идеология данного подхода базируется на возрождении типологической концепции вида и макромутационистском толковании проблемы видообразования. Убедительным аргументом этого являются геномные мутации и, прежде всего, полиплоидия. Около 70% видов покрытосеменных растений возникло путем полиплоидии - резкого скачкообразного преобразования видового генома [12]. Этот способ видообразования, кроме растений, отмечен и в разных группах животных и является единственным в природе доказательным (и воспроизводимым человеком) примером сальтационного видообразования. Хорошо увязываются с сальтационным принципом видообразования и скачкообразные модификации гетерохроматиновых компонентов генома. К настоящему времени получено много данных относительно реорганизации гетерохроматиновых блоков хромосом в близкородственных видовых комплексах. Основной закономерностью при этом является локализация гетерохроматина в прицентромерных блоках и в микросомах у «стволовых» видов и «диспергирование» гетерохроматина по хромосомным плечам у видов, занимающих терминальные положения в филогенетических ветвях. С сальтационным видообразованием хорошо согласуются и данные по мобильным генетическим элементам, обнаруживающим видовую специфику [10, 13].
Все известные с начала XX века мутации были подразделены на три типа: генные, хромосомные и геномные. К настоящему времени стало очевидным, что существует еще один тип — системные мутации.
В 1979 г. был выявлен феномен кардинальной реорганизации архитектуры интерфазных ядер в онто- и филогенезе малярийных комаров |8|, затем подтвержденный на дрозофиле [14, 15]. Именно анализ архитектуры ядер генеративных клеток позволил нам выявить наиболее существенные закономерности: 1) принципиальное различие архитектуры хромосом в клетках генеративной ткани и в соматических клетках; 2) существенные различия в трехмерной организации хромосом в самих генеративных клетках между разными (в том числе и очень близкими) видами.
Видоспецифичность проявляется в следующих показателях: 1) наличие – отсутствие связей хромосом с ядерной оболочкой и локализация мест контактов на хромосомах; 2) морфология хромосомных участков прикрепления; 3) разобщенность локусов прикрепления гомеологичных хромосом у близких видов на ядерной оболочке; 4) наличие – отсутствие локального или диффузного хромоцентра.
Взаимоотношения хромосом с ядерной мембраной облигатно инвариантны в пределах каждого вида малярийных комаров и четко различаются между восемью изученными видами (рис. 1). У межвидовых гибридов проявляются видоспецифичные особенности каждого из видов, причем значительная пространственная разобщенность зон прикрепления каждого из гомеологов в гибридных клетках свидетельствует о видоспецифичности координат точек прикрепления на мембране.
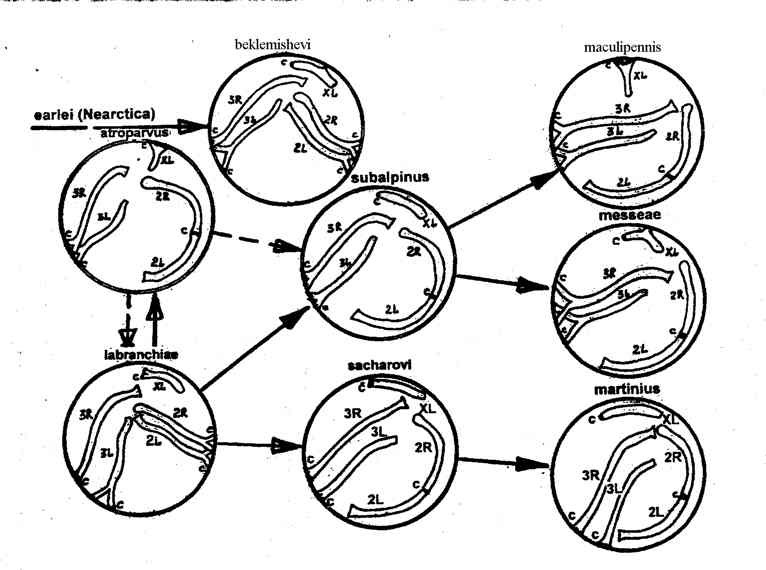
Рис.1. Схема эволюционного изменения архитектуры политенных хромосом трофоцитов яичников восьми видов малярийных комаров палеарктического комплекса Anopheles maculipennis. Стрелками показаны направления видообразования (сплошными - наиболее вероятные, пунктирными - менее вероятные).
На основании данных по видовым комплексам "D. melanogaster" и "D. virilis" можно оценить основные закономерности реорганизации хромосомной архитектуры в филогенетических направлениях от стволовых (исходных) видов к терминальным (производным) видам [14, 15]. Общая схема такова: у стволовых видов (D. orena, D. virilis, D. kanekoi, D. flavomontana) наблюдается концентрация гетерохроматина в области хромоцентра, затем у производных видов хромоцентр визуально исчезает, и хромосомы демонстрируют либо свободное (без прикрепления к ядерной оболочке) состояние (D. mauritiana, D. teissieri, D. yakuba), либо частичное (отдельные хромосомы) прикрепление (D .novamexicana) (рис. 2).

Рис.2. Схема эволюционного изменения архитектуры первично политенных хромосом трофоцитов яичников восьми видов комплекса Drosophila melanogaster. Стрелками показаны направления видообразования, основанные на комплексных данных [14]
Обнаруженный принцип хромосомной реорганизации рассматривается нами как явление мутационного ранга, не имеющего аналогов среди известных типов хромосомных мутаций. Таким образом, перестройку архитектуры интерфазного ядра можно оценить как новый тип мутаций, названный нами системными мутациями.
Системные мутации возникают в результате пространственной реорганизации интерфазных хромосом в ядре за счет изменения хромосомно-мембранных взаимоотношений. Системные мутации в своем происхождении связаны с перестройкой хромоцентрального аппарата: изменяется структура хромоцентра (от локального до диффузного) вплоть до визуального исчезновения последнего; появляется (или исчезает) прикрепление центромерных, теломерных и других хромосомных локусов к ядерной оболочке. Особенностью системных мутаций является также то, что они: 1) четко выявляются только в клетках генеративной ткани; 2) являются видоспецифичными и не обнаруживают внутривидового полиморфизма; 3) проявляют «гетерозиготность» только у межвидовых гибридов, геномы которых отражают различия в архитектуре хромосом «отцовского» и «материнского» видов.
Таким образом, архитектура хромосомного аппарата генеративной ткани представляет собой элементарную типологическую систему, отражающую видоспецифичность признаков.
Происхождение системных мутаций. Как выше было отмечено, перестройка архитектуры хромосомного аппарата, или системные мутации, были обнаружены только в питающих клетках (трофоцитах) яичников самок комаров и дрозофилы. Изучение соматических тканей с политенными хромосомами (слюнные железы, мальпигиевы сосуды) не выявило каких-либо очевидных различий в пространственной организации хромосом между видами. Для всех изученных видов малярийных комаров в соматических тканях существует универсальная организация хромосом: они объединены центромерными районами в единый хромоцентральный узел по принципу эктопической конъюгации. Для дрозофил также характерна однотипная система организации политенных хромосом слюнных желез, хотя и хромоцентральное объединение центромерных участков представляет более диффузное хроматиновое «облако», чем у комаров. Интересно, что у межвидовых гибридов F1 в клетках слюнных желез малярийных комаров хромоцентр остается общим для гомеологов «отцовского» и «материнского» видов при отсутствии конъюгации между гомеологичными хромосомами (частичном — у аутосом и полном — у половых хромосом).
Таким образом, учитывая происхождение питающих клеток, системные мутации относятся непосредственно к генеративной сфере. Действительно, исследования, проводимые уже более 80 лет (Э. Вильсон, X. Равен, R. King), убедительно и однозначно свидетельствуют, что трофоциты у организмов с нутриментарным (алиментарным) типом оогенеза являются сестринскими с собственно ооцитом. В генеративной ткани самок первичные оогонии делятся несколько раз (у малярийных комаров - 3 раза, у дрозофилы - 4 раза). В результате возникает 8 клеток у комаров и 16 - у дрозофилы. Центральная по суперпозиции клетка становится ооцитом, остальные (7 - у комаров и 15 - у дрозофилы) - питающими (трофоцитами), R. King [16] показал, что ооцит связан со всеми остальными сестринскими клетками (трофоцитами) цитоплазматическими мостиками. Впоследствии ооцит претерпевает мейотические преобразования, а питающие клетки остаются диплоидными. В начале периода созревания фолликула хромосомы трофоцитов интенсивно политенизируются и с высочайшей активностью синтезируют РНК-продукты, практически полностью обеспечивая рост и развитие ооплазмы, и, что самое главное, формируют ооплазматическую сегрегацию.
Системные мутации наиболее четко проявляются в период максимального развития политенных хромосом у малярийных комаров. У дрозофилы в норме в трофоцитах существуют так называемые первично-политенные хромосомы, размеры которых значительно меньше таковых у комаров, и между тем системные мутации проявляются достаточно четко.
Методически пока невозможно оценить архитектуру хромосом в оогониях и ооците. Однако есть все основания считать, что топологические параметры взаиморасположения отдельных хромосом и их связи с ядерной оболочкой в оогониях и ооците так же, как и у трофоцитов, носят видоспецифичный характер. Наши данные показывают, что для ооцита малярийных комаров характерно отсутствие общего локального хромоцентра, что совпадает с данными по трофоцитам, следовательно, схема хромосомной организации у них идентична. В пользу этого свидетельствуют также «гибридные конфигурации» (F1) ядер ооцитов на стадии пахитены (от скрещивания Anopheles maculipennis и A. subalpinus), показывающие отсутствие конъюгации гомеологов хромосом, как и в «гибридных» трофоцитах [9].
В заключение, касаясь принципов пространственной организапии хромосом половых клеток (яйцеклеток и спермиев), необходимо отметить, что методические ограничения визуализации их хромосом не позволяют пока детально оценить характер топологии. Было бы опрометчиво предполагать, что архитектура хромосом и трофоцитов идентична, поскольку и структурно и функционально эти системы различаются, хотя и имеют общее происхождение. Однако есть достаточно оснований считать, что и в ооците, и в оогониях, а также в спермиях и сперматогониях элементы видоспецифичной архитектуры хромосом идентичны трофоцитам. В этом отношении важные данные демонстрируют межвидовые гибриды малярийных комаров. В трофоцитах самок F1 обнаруживаются все специфичные элементы организации хромосом, характерные для каждого из родительских типов. Впечатляет то, что посредством спермиев от родителя-самца передается полная информация относительно видоспецифичной архитектуры хромосом в трофоцитах самок своего вида. Это достаточно убедительный аргумент в пользу видоспецифичности хромосомной архитектуры ядер в генеративной ткани в целом с подобными же видоспецифичными особенностями в отдельных клеточных системах ткани. Клетки зародышевого пути от оогониев в гермарии до зрелого ооцита (у самок) и от сперматогониев до сперматозоидов (у самцов) определенно имеют видовую специфику в структурной организации ядерного материала, как это предполагал известный цитолог конца XIX века О. Гертвиг.
В следующей статье предполагается осветить взаимодействие видовой системы с внешней средой и прежде всего вскрыть механизмы трансформации видового генома в видообразовательном смысле.
1 Голубовский М.Д Организация генотипа и формы наследственной изменчивости
эукариот // Успехи совр. биологии 1985 Т. 100 Вып. 6. С. 323-339.
2. Садовский В.Н. Основания обшей теории систем. М Наука, 1974. С. 274.
3. Алтухов Ю.П.. Рычков Ю.Г. Популяционные системы и их структурные компоненты. Генетическая стабильность и изменчивость // Журн.общ.биологии. 1970. Т 31. № 5 С. 507-526.
4. Майр Э Зоологический вид и эволюция. М.: Мир, 1968. 597 с.
5. Dobzhansky Th. Genetics of evolutionary process. NY.,L. Columbia Univ. Press, 1971. 520 p.
6. Стегний В.Н. Популяционная генетика и эволюция малярийных комаров. Томск. Изд-во Том. ун-та, 1991. 137 с.
7 Алтухов Ю П., Рычков Ю.Г. Генетический мономорфизм видов и его возможное биологическое значение // Журн общ. биологии. 1972. Т. 33 № 3. С. 281-300.
8 Стегний В.Н. Реорганизация структуры интерфазных ядер в онто- и филогенезе малярийных комаров // ДАН СССР. 1979 Т. 249 № 5. С. 1231
9. Стегний В.Н. Архитектоника генома, системные мутации и эволюция. Новосибирск: Изд-во Новосиб. ун-та. 1993. 110с
10. Евгеньев М.Б., Ениколопов Н.П.. Пеунова Н.И. Транспозиция мобильных диспергированных генетических элементов у дрозофилы // ДАН СССР 1982 Т 64. № 6. С 145.
11. Тахтаджян А.Л. Макроэволюциоиные процессы в истории растительного мира
// Ботан журн. 1983. Т. 68. № 12. С. 1593 -1603.
12. Грант В. Видообразование у растений. М.: Мир, 1980. 407 с
13. Корочкин Л.И. Эволюционное значение генетических подвижных элементов // Цитология и генетика. 1983. Т. 17. № 4. С. 67-78.
14. Стегний В.Н., Вассерлауф И.Э. Видовая архитектоника хромосом генеративной ткани и проблемы филогенетических отношений в подгруппе melanogaster рода Drosophila ( Sophophora) // Генетика. 1994. Т. 30. № 4. С. 478-483.
15 Стегний В.Н , Вассерлауф И.Э., Ананьина Т.В. Взаиморасположение первичных политенных хромосом яичников у 12 видов группы «virilis» рода Drosophila (Sophophora) // Генетика 1996. Т. 32. № 6. С. 750-754.
16. King R. The meiotic behavior of the Drosophila oocyte// Intern Rev. Cytol. 1970. Vol. 28. P 125.