
Проблемы Эволюции |
Диссертация. Москва, 2002.
Дмитриев Виктор Юрьевич
Москва
2002
Актуальность темы. Разнообразие ископаемых организмов и его изменения во времени - один из наиболее общих вопросов палеонтологии, требующий привлечения обширного эмпирического материала. Хотя проблема относится к числу классических, обсуждение методов ее решения не проведено с необходимой полнотой. В связи с этим не до конца прояснены вопросы ограничений в использовании того или иного фактического материала, в интерпретации результатов его изучения и техники исследований.
Цели и задачи. Целью данного исследования было обоснование рациональных подходов к описанию эволюции таксономического разнооб-разия и выяснение на этой основе главных особенностей диверсификации организмов прошлого.
Материал. В методической части работы для иллюстрации основных положений был привлечен материал по родам и семействам морских животных. Сведения по родам были взяты из неопубликованной базы дан-ных Дж. Сепкоски, переданной им в Палеонтологический институт РАН незадолго до смерти и содержащей датировки первых и последних находок для 35638 родов. Данные по семействам были опубликованы Дж. Сепкоски ранее (Sepkoski, 1982, 1992). Использованы также сведения по се-мействам насекомых, собранные коллективом палеоэнтомологов Палеон-тологического института РАН, в сравнении с материалами, опубликован-ными независимо К. Лабандейра (Labandeira, 1994). При изучении вопроса о полноте ископаемого материала анализировались данные по родам морс-ких гастропод позднего мела - палеогена (403 рода), опубликованные кол-лективом авторов в сводке “Развитие и смена моллюсков на рубеже мезо-зоя и кайнозоя” (1981). Для изучения эволюции континентального биораз-нообразия использованы данные по пресноводным животным (915 се-мейств), тетраподам (931 семейство) и насекомым (1049 семейств). Иссле-дование континентальной биоты проводилось по инициативе А.Г. Понома-ренко, который получил необходимый материал от большого коллектива специалистов и передал его автору для обработки. В подготовке данных участвовали В. А. Алифанов, А. Ф. Банников, В. А. Благодеров, А. В. Го-рохов, Ю. М. Губин, К. Ю. Еськов, В. В. Жерихин, М. Ф. Ивахненко, Н. Н. Каландадзе, Е. Н. Курочкин, О. А. Лебедев, Е. Д. Лукашевич, В. Г. Новок-шенов, А. Г. Пономаренко, Ю. А. Попов, Л. Н. Притыкина, А. П. Расни-цын, А. С. Раутиан, А. Г. Сенников, Н. Д. Синиченкова, Я. И. Старобога-тов, И. Д. Сукачева, В. Б. Суханов, М. А. Шишкин, Д. Е. Щербаков.
Защищаемые положения:
1. Наблюдаемые скорости появления и вымирания родов и семейств морс-ких животных, а также скорости появления семейств пресноводных жи-вотных и насекомых в течение длительных интервалов времени (100 - 250 млн. лет и более) вариируют около некоторых постоянных.
2. Минимумы разнообразия не всегда обусловлены увеличением скорости вымирания. До выяснения кинематических характеристик процесса в ок-рестностях минимума не следует интерпретировать минимум как массовое вымирание.
3. Выявлено коллективное поведение приращений разнообразия (разность числа появившихся и вымерших родов) в разных группах морских живот-ных.
4. Основные минимумы разнообразия морских, пресноводных и наземных животных совпадают.
5. Актуальное (наблюдаемое, опубликованное) разнообразие и потенциальное (доступное) разнообразие, выделенные ранее Н. Ньюеллом (Newell, 1959), следует дополнить еще двумя категориями - полное (захоронившееся) и общее (существовавшее) разнообразия. Основным вопросом эмпири-ческого изучения диверсификации ископаемых организмов является со-отношение актуального и полного разнообразия.
6. Для адекватного отображения исходных данных используются графики мгновенного числа таксонов, показывающие результаты соизмерения мгновенных морфологических разнообразий. Применение любых выбо-рочных показателей нельзя считать оправданным, как основанное на апри-орном признании актуального разнообразия статистически репрезентатив-ной выборкой.
7. Зависимость время - число появившихся родов можно использовать в геохронометрии.
Научная новизна. Тезисы 1, 3-7 “Защищаемых положений”- новые, тезис 2 - новый в части диагностики массовых вымираний.
Теоретическая значимость работы. Изучение скоростей появления и вымирания семейств и родов морских и континентальных животных ука-зывает на единство биоты в процессе ее диверсификации с признаками са-моорганизации и саморегуляции. Пересмотрены и дополнены общие пред-ставления о минимумах разнообразия и массовых вымираниях. Предло-женный метод детализации изотопно-геохронологической шкалы на осно-ве палеонтологических данных может быть перспективен для решения различных проблем геологии и палеонтологии.
Аппробация работы. Основные положения диссертации докладыва-лись на коллоквиумах Лабораторий брахиопод и членистоногих ПИН (1976, 1998), школе-семинаре “Математика в палеонтологии” (Кишинев, 1978), совещаниях “Редкие события в геологии” (Ашхабад, 1985), “Таксо-номическое разнообразие: систематическая изученность таксонов органи-ческого мира и формирование компьютерных банков данных” (Москва, 1991), “Эволюция биосферы” (Абрау-Дюрсо, 1990; Москва, ПИН, 1998, 1999, 2000), Методологическом семинаре ПИН (1988), сессии Палеонтоло-гического общества (Санкт-Петербург, 2001). По теме диссертации имеется 12 публикаций.
Структура работы. По своей структуре диссертация в основном следует изложению материала в книге: Алексеев А.С., Дмитриев В.Ю., По-номаренко А.Г. Эволюция таксономического разнообразия. М.: ГЕОС, 2001, где автором написаны главы “Кривые разнообразия” (с. 19-67) и “Палеонтологическая интерполяция в геохронометрии” (с. 111-118), а так-же совместно с А.Г. Пономаренко глава “Эволюция разнообразия конти-нентальных биот” (с. 68-95). Первая из них посвящена преимущественно принципиальным основам изучения диверсификации древних организмов. Содержит также обзор основных результатов по морским животным. В главе о континентальной биоте приведены данные по эволюции разнооб-разия пресноводных животных, насекомых и тетрапод на уровне семейств. Третья глава посвящена использованию палеонтологических данных для совершенствования геохронометрической шкалы. В диссертации отдель-ные положения указанной монографии уточнены, в основном во второй главе.
Благодарности. Автор благодарен за поддержку данной работы Г.А. Афанасьевой, М.В. Базилевскому, Л.Н. Большаковой, М.К. Емельяновой, В.В. Жерихину, И.В. Кирилловой, С.С. Лазареву, М.Б. Мостовскому, Л.А. Невесской, А.Г. Пономаренко, А.П. Расницыну, А.Ю. Розанову, а также всем перечисленным выше специалистам, участвовавшим в подготовке данных по континентальной биоте. Автор особо признателен Дж. Сепкос-ки за материал по морским животным.
В палеонтологии традиционно используется описание эволюции биологического разнообразия на основе изменений во времени числа так-сонов того или иного ранга. Это связано с доступностью исходных данных не только для отдельных крупных групп организмов, но и всех групп вмес-те (по крайней мере морских), особенно после появления многотомных мировых справочников и специальных списков. В отличие от числа таксо-нов, изменения экологического, морфологического, генетического разно-образия, числа сообществ, частотных (информационных) мер и т. п., как правило, изучаются ограниченно - для отдельных групп организмов, ло-кальных местонахождений или групп местонахождений, сравнительно не-больших временных интервалов. Это определяется сложностью и неодно-значностью, во-первых, подбора достаточно однородного исходного мате-риала и, во-вторых, его численного выражения, что влечет за собой труд-ности при интерпретации получаемых результатов. Более подробные ис-следования имеются по видовому богатству древних сообществ (Bambach, 1986).
Термин “разнообразие” в современной фаунистике применяется для обозначения некоторых количественных характеристик, зависящих от чис-ленности видов. В палеонтологии “разнообразие” - синоним числа таксо-нов того или иного ранга. Это различие в словоупотреблении является ис-точником недовольства со стороны некоторых пуристски ориентирован-ных биологов, но не приводит к большим недоразумениям. “Эволюция разнообразия” в палеонтологии - привычные, хорошо укоренившиеся сло-ва, а палеонтологическая литература, относящаяся к этой области исследо-ваний, достаточно изолирована от неонтологической литературы по фау-нистике.
Хорошо известны два фундаментальных факта: сравнение кривых разнообразия крупной группы при использовании разных вариантов ее системы обнаруживает независимость (или слабую зависимость) результа-тов от системы группы (Maxwell, Benton, 1990). Точно так же, общая кар-тина изменений разнообразия обнаруживает замечательное постоянство в процессе изучения: повторные исследования через много лет после преды-дущих показывают в общем те же особенности (Sepkoski et al., 1981; Sep-koski, 1993b). Число таксонов одного ранга в исходном фактическом мате-риале и данные об их распространении сильно меняются, но форма кривой разнообразия устойчиво сохраняется. Отсюда естественно вытекает пред-ставление о том, что числом таксонов измеряется некоторая постоянная для каждого момента геологического времени величина, и возникает воп-рос о ее природе.
Он решается на основе введенного Н. Ньюеллом (Newell, 1959) по-нятия о потенциальном разнообразии, подразумевающего все доступное полноценному палеонтологическому изучению. Все уже изученное, то есть пригодное для непосредственного подсчета и реально используемое (актуальное разнообразие), составляет лишь часть потенциального разно-образия. Устойчивость кривых разнообразия свидетельствует о достаточно близком соответствии актуального и потенциального разнообразия, но связанные с этим вопросы нуждаются в специальном обсуждении.
Измерение актуального и даже потенциального разнообразия не ох-ватывает всего захороненного в прошлом разнообразия остатков организ-мов (полное разнообразие). Поэтому возникает необходимость оценки и учета появляющихся из-за этого искажений. Точно также полное разно-образие есть только часть всего существовавшего (или общего) разнообра-зия. Последнее находится вне палеонтологических исследований, которые могут дать материал только для некоторых гипотетических предположе-ний о его изменениях во времени. Данная глава ограничивается, поэтому, соотношениями актуального, потенциального и полного разнообразий, не-доучет которых может приводить к появлению артефактов.
Палеонтологическое “измерение” разнообразия отличается от обыч-ных измерений в других дисциплинах. Для каждого момента геологичес-кого времени число одноранговых таксонов увеличивается как в результа-те находок неизвестных ранее ископаемых организмо, так и ревизии мор-фологии и системы групп. Единица измерения постоянно меняется. Это вынуждает несколько подробнее остановиться на вопросе о таксонах как единице измерения разнообразия и приводит к обсуждению некоторых возможностей перехода к измерению разнообразия без таксонов.
Со словом “измерение” неизбежно связан вопрос о его точности, то есть полном перечне источников погрешностей и способов их оценки. Собственно палеонтологические погрешности связаны с искаженным ото-бражением полного разнообразия в актуальном и с измерением разнообра-зия числом таксонов. Погрешности, возникающие вследствие несовершен-ства изотопно-геохронологической шкалы и стратиграфической корреля-ции, также должны учитываться.
Другая интерпретация устойчивости кривых разнообразия основана на коллективном поведении основных групп морских животных и не нуж-дается в представлении об измерении. Она не альтернативна первой интер-претации и совместима с ней, хотя дает иное толкование устойчивости кривых. Основное внимание в данной главе уделяется первой интерпрета-ции как более перспективной в связи с более широкими возможностями ее применения.
Данная работа была инициирована А. Г. Пономаренко и изначально планировалась как преимущественно методическая по своей направленно-сти. Она уже составлялась, когда были получены материалы Дж. Сепкоски по родам морских животных. Их использование чрезвычайно расширило возможности обсуждения, и в результате методическая по замыслу глава построена по классической схеме: методика - результаты - обсуждение. Методике, точнее технике первичной обработки данных посвящен краткий раздел, затем рассмотрены результаты по морским животным и менее подробно - по семействам насекомых. Раздел по морским животным не имеет своей целью полное прояснение происходивших событий, а толь-ко указывает на некоторый круг возможностей. В обсуждении анализиру-ются принципы, лежащие в основе кривых разнообразия, и ограничения метода. Наиболее существенное из ограничений связано с неравномерной и неполной палеонтологической изученностью групп и фаун. Это частич-но компенсируется повышением ранга таксонов, что приводит к огрубле-нию результатов.
Число таксонов N(t) в каждый момент времени t определяется балан-сом появления новых таксонов N+(t) и вымирания старых N-(t) за все время до момента t:
N(t) = N+(t) - N-(t) (1)
Имитационные модели процесса могут создаваться на основе уравнений для скоростей
dN+/dt = l(t)
dN-/dt = m(t) (2)
или, если предполагается зависимость скоростей от числа таксонов N,
dN+/dt = l(N)
dN-/dt = m(N). (2a)
Такие модели легко конструируются, если ввести явные выражения для l(t), m(t) или l(N), m(N), исходя из каких-либо простых теоретических предпосылок или эмпирических данных; балансовое соотношение (1) рас-сматривается тогда как описывающее математические ожидания в некото-ром ветвящемся случайном процессе. Уравнения (2а) совпадают с приме-няемыми в экологии для описания численности изолированной популя-ции. Имитационные модели применяются лишь изредка, когда удается предположительно указать явный вид скоростей (Дмитриев, 1978), обычно же ограничиваются построением и описанием графиков.
Поскольку основные уравнения связывают мгновенные значения пе-ременных, графики также должны показывать мгновенные величины. Тра-диционно многие палеонтологи используют число таксонов N(t), сущест-вовавших в течение века, то есть по оси ординат откладывается сумма мгновенного числа таксонов в начале века N0 и числа появившихся в этом веке таксонов DN+, за абсциссу принимается середина века, что описывает-ся равенством
N(t) = N+(t+t) - N-(t),
t - продолжительность века. Наклон такого графика между смежными точ-ками определяется равенством
tga = (DN+2 - DN-1)/(t2 - t1),
где t2 - t1 - временной интервал межде серединами веков, DN-1 - число так-сонов, вымерших в более древнем веке, DN+2 - число таксонов, появивших-ся в более молодом веке. График верно отражает локальное возрастание или убывание разнообразия, только если DN+2 и DN-1 случайно оказались подходящими (что бывает нередко), его наклон вообще не зависит от при-ращения разнообразия в течение века. Хотя уже давно в некоторых публи-кациях использовалось мгновенное разнообразие (Webb, 1969), во многих современных исследованиях продолжается применение некорректного об-щего числа таксонов N(t). Традиционные кривые могут мало отличаться своей общей формой от кривых, построенных на основе мгновенного чис-ла таксонов (если взяты малые временные интервалы), но они полностью исключают возможность локального анализа кривых разнообразия.
Исходные палеонтолого-стратиграфические данные не позволяют непосредствено выяснить число таксонов для любого момента. Единствен-ный способ эмпирического определения мгновенных величин - подсчет для границ геохронологической шкалы (Дмитриев, 1978, 1989, 2001; По-номаренко, Дмитриев, 1993; Foote, 2000) равенство (1) конкретизируется тогда следующим образом:
N(ti) = N(to) + N+(ti) - N-(ti),
где ti - возраст границы с номером i; N(to) - число таксонов, существовав-ших в начале изучаемого возрастного интервала; N(ti) - число таксонов, су-ществовавших на границе с номером i (то есть общих для смежных под-разделений); N+(ti) - число таксонов, появившихся в интервале от to до ti; N-(ti) - число таксонов, вымерших в интервале от to до ti. Предлагалось ис-пользовать среднее арифметическое числа таксонов на границах и отно-сить его к середине подразделений (метрика Вебба-Харпера; Webb, 1969; Harper, 1975). Эта величина дает оценку среднего мгновенного разнообра-зия в течение века и именно так применяется ниже. Она может служить также оценкой мгновенного разнообразия в середине века, но ее использо-вание в этом качестве для отображения изменений разнообразия представ-ляется излишней данью традиции.
Для полной характеристики процесса нужны два графика. График N(t) иллюстрирует результат процесса, но для вскрытия его структуры (соотношения появления и вымирания таксонов) требуется еще один, на-пример N+(t). Всю необходимую информацию содержит любая пара вели-чин в равенстве (1), либо произведенных от них, как скорости или удель-ные скорости. На практике выбор графиков определяется целью конкрет-ного исследования и необходимостью обеспечить более доступное воспри-ятие. Последнее часто вынуждает использовать более двух графиков. Яс-ное и наглядное представление об изучаемом процессе дает совместный анализ трех графиков: N+(t), N- (t), N(t) .
Погрешности величин N, N+, N- зависят только от качества палеон-толого-стратиграфических данных, хотя на наклон графиков N(t), N+(t), N- (t) между соседними точками влияют также ошибки в определении про-должительности веков, которые могут быть очень велики, особенно до позднего мела (Рублев, 1996). Графики N+(t), N-(t) палеонтологами не ис-пользуются, а изучаются скорости или удельные скорости вымирания и появления таксонов. Вследствие ошибок продолжительностей оценки ско-ростей могут быть слишком грубыми, поэтому лучше изучать непосредст-венно N+(t), N-(t). Скорости отражаются тогда наклоном графика, причем зрительно воспринимаемый угол наклона сравнительно мало чувствителен к их изменениям. Это в данном случае оказывается удобным, так как оцен-ки самих скоростей недостаточно надежны.
Погрешности скоростей в значительной мере сглаживаются с увели-чением временных интервалов: относительная погрешность продолжи-тельностей двух-трех или нескольких смежных веков вместе меньше. При отсутствии резких хаотичных изменений скоростей усредненный наклон графика на ряде смежных временных интервалов дает более адекватное представление о скоростях и долговременных тенденциях их изменений. Как будет проиллюстрировано на конкретном материале ниже, все же кро-ме графиков N+(t), N-(t) приходится использовать и оценки скоростей для отдельных веков.
Другой тип графиков использует координаты (N+,N) (Дмитриев, 1978, 1989), что исключает влияние погрешностей временной шкалы и обычно упрощает форму кривой. График позволяет детализировать изуче-ние процесса, так как не зависит от изотопно-геохронологической шкалы. Функция N(N+) полезна еще в одном отношении (Дмитриев, 1978). Если предпринимается попытка описать процесс на основе уравнений (2) или (2а), необходимо сделать два предположения о скоростях. Но если зависи-мость N(N+) установлена эмпирически, достаточно одного предположения, так как
dN/dN+ = 1 - m(t)/l(t).
Это обеспечивает возможность исследовать гипотезу об одной из скорос-тей. Если гипотеза подтверждается, описание второй скорости может быть получено как следствие. Точно так же можно использовать зависимость N(N-) или N+(N-) - это вопрос удобства, решаемый применительно к осо-бенностям изучаемого материала (для аппроксимации выбирается более простая из трех кривых).
Можно дополнительно отметить некоторые частные особенности графиков N(t), N(N+). Если число таксонов в течение века не изменяется, на графиках первого типа точки на его границах располагаются на концах горизонтального отрезка. На графиках второго типа то же будет, если в те-чение века установлено появление и вымирание таксонов. В противном случае точки для обеих границ совпадают, и на графике вместо двух ока-зывается только одна точка. Тем самым на графиках N(N+) наглядно разли-чаются века сбалансированного появления и вымирания таксонов и века отсутствия того и другого, что не выявляется при использовании только графиков N(t). Эволюционное различие этих двух ситуаций вполне оче-видно, но здесь необходима определенная осторожность, так как нулевой случай может быть и следствием слабой изученности фауны соответству-ющих отложений. Наклон графиков N(N+) зависит только от отношения числа появившихся и вымерших таксонов - важной характеристики хода диверсификации изучаемых групп или фаун. При закономерных изменени-ях или постоянстве этого отношения такие графики выявляют это легко и наглядно.
Оба типа графиков с ординатой N не учитывают (как составную часть мгновенных разнообразий) таксоны, известные только из отложений одного века. Такие таксоны бывают двух типов. Первый, это действитель-но кратковременно существовавшие таксоны, распространение которых в пределах века известно. Их вклад в изменения разнообразия может быть изучен при детализации используемой хронологической шкалы, то есть повышении ее временного разрешения. Второй тип таксонов составляют уникальные находки из локальных местонахождений. Эти таксоны имеют только морфологическую характеристику, время их появления и вымира-ния не установлено, то есть отсутствует характеристика распространения, а известное время существования равно 0. Процедура обработки данных автоматически отказывается включать такие таксоны в состав мгновенных разнообразий (так же как они не могут быть составной частью материала для построения кривых выживания). Эта проблема относится к области полноты палеонтологической изученности и устраняется только при ее по-вышении. При полной палеонтологической изученности и достаточной де-тальности хронологической шкалы вопрос об учете всех таксонов, пред-ставители которых присутствуют в тех или иных отложениях, решается сам собой, но лишь в перспективе дальнейших исследований.
На графиках N+(t), N-(t) таксоны, описанные по уникальному мате-риалу, отображаются, хотя правильнее исключать их при подготовке дан-ных (Sepkoski, 1996; Foote, 2000; Марков, 2001, 2002), оставляя для анали-за короткоживущие таксоны с установленным распространением в преде-лах века. В перечисленных публикациях исключаются все роды, появив-шиеся и вымершие в одном и том же интервале (веке или его части), что не вполне точно (полностью теряются значительные части таких характер-ных комплексов как хирнантиевая фауна в конце ордовика, а при дробле-нии шкалы только до веков также специфические фаменские представите-ли - отряд Clymeniida и др. или альбские Hoplitaceae). В данной работе ис-ключение уникального материала не делалось, так как исходные списки не содержат сведений, необходимых для отдельного учета только таксонов, распространение которых не охарактеризовано. Ниже, на примере ископа-емых насекомых частично иллюстрируется влияние этого фактора.
Исходные данные подвергались минимальной предварительной об-работке, направленной на использование числа появлений или вымираний таксонов, указанных интервалом возрастов. Если таксонов с такими дати-ровками оказывалось меньше, чем точно датированных в этом же возраст-ном интервале, то число появлений или вымираний распределялось по наиболее дробным хронологическим интервалам пропорционально числу точно установленных в них появлений или вымираний (Дмитриев, 1989; Sepkoski, Koch, 1996). В противном случае дробные интервалы объединя-лись и соответствующие числа суммировались. Тем самым особенности фактического материала определяют возможную в данном исследовании детальность хронологической шкалы, которая формируется автоматически на основе простейшего алгоритма.
Ниже приводятся результаты изучения морских животных и се-мейств насекомых. Исходным материалом по морским животным послу-жили данные Дж. Сепкоски: опубликованный список по отрядам и се-мействам (Sepkoski, 1982) и переданный им в Палеонтологический инсти-тут РАН список родов. Для насекомых приведено сравнение результатов обработки двух независимо составленных списков семейств - Лаборатории членистоногих ПИН и список К. Лабандейра (Labandeira, 1994). Во всех случаях использовалась временная шкала Американского геологического общества, 1989 г. (Geologic Time Scale, Decade of North American Geology, 1980-1989. Geol. Soc. Amer., 1989).
Сопоставление кривых на разных таксономических уровнях. На рис. 1 показаны кривые числа отрядов, семейств и родов морских живот-ных (387 отрядов, 3716 семейств, 35638 родов) от томмотского века до современности, приведенные к единой шкале делением на число совре-менных таксонов, известных в ископаемом состоянии (166 отрядов, 1034 семейств, 5155 родов). Наиболее подробной является кривая для родов, так как датировки родов более детальны, чем семейств и отрядов, а объе-динение смежных веков производилось реже. Кривые обнаруживают вы-сокую степень сходства и ряд характерных отличий, обсуждаемых ниже.
<![if !vml]>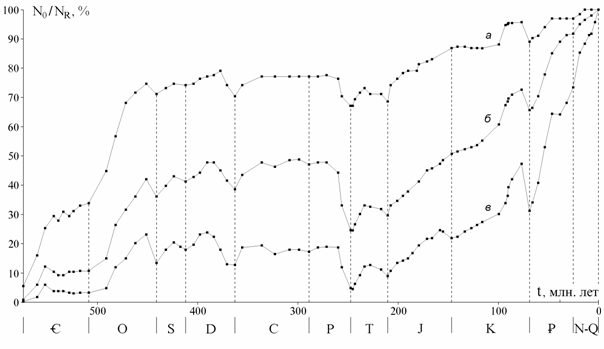 <![endif]>
<![endif]>
Рис. 1. Изменения разнообразия морских животных: а - отряды, б - семейства, в - роды. N0 - число таксонов, существовавших на нижней границе века, NR - число сов-ременных таксонов, известных в ископаемом состоянии. Данные для рис. 1 - 3, 10: от-ряды и семейства - по Sepkoski, 1982; роды (также на рис. 4 - 8, 10) - из неопубликован-ного списка Сепкоски, переданного в Палеонтологический ин-т РАН
Сходство можно измерить с помощью коэффициента корреляции, но при этом нужно учесть различия их общей конфигурации: кривая отрядов в целом выпуклая, родов - вогнутая, семейств - также вогнутая, но в мень-шей степени и занимает промежуточное положение между двумя другими. Коэффициент прямолинейной корреляции здесь неприменим, но можно учесть обращающее на себя внимание сходство всех трех кривых в палео-зойской и в мезокайнозойской их частях. Все коэффициенты корреляции кривых отдельно для палеозоя и мезокайнозоя оказываются в пределах 0.94 - 0.98, за исключением коэффициента для мезокайнозойской части кривых по отрядам и родам равного 0.85.
Связь числа таксонов разного ранга. Сравнение общей формы кривых для родов, семейств и отрядов оставляет впечатление возможности легко найти уравнение регрессии. Действительно, связь числа таксонов смежных рангов приблизительно описывается зависимостью (рис. 2)
Ni+1 = A(lnNi) r, (3)
где Ni - число таксонов, Ni+1 - число таксонов следующего более высокого ранга, A, r - параметры. Параметры для родов и семейств (рис. 2, а): A = 0.2035, r = 4.000, коэффициент корреляции фактического и вычислен-ного числа семейств 0.98. Для семейств и отрядов (рис. 2, б): A = 1.310,
<![if !vml]> <![endif]>
<![endif]>
Рис. 2. Сопоставление фактического (квадратики) и вычисленного (крестики) числа отрядов и семейств: а, б - по уравнению (3); а - семейства в зависимости от чис-ла родов, б - отряды в зависимости от числа семейств; в - отряды в зависимости от чис-ла родов по уравнению (4)
r = 2.528, коэффициент корреляции также 0.98. Связь числа родов и отря- дов (рис. 2, в) определяется непосредственно (без оценки параметров) из двух предыдущих зависимостей
Ni+2 = 1.310(4.000lnlnNi - 1.592)2.528 (4) Форма кривой числа отрядов и в этом случае воспроизводится удовлетво-рительно. Коэффициент корреляции равен 0.93 (корреляция полных кри-вых для родов и отрядов на рис. 1 равна 0.68). Наибольшие отличия факти-ческих и вычисленных кривых при всех трех сопоставлениях относятся к их кембрийской части и пермо-триасовому минимуму разнообразия. Воз-можность однотипного преобразования кривых для всего фанерозоя пред-
<![if !vml]> <![endif]>
<![endif]>
Рис. 3. Число появившихся и вымерших семейств и родов: а, б - семейства; а – появившиеся, б - вымершие; в, г - роды; в - появившиеся, г - вымершие. Данные по ар-хеоциатам не учитывались, то же на рис. 5-8 и 10
ставляет несомненный интерес и нуждается в теоретическом истолкова-нии.
Фоновые скорости появления и вымирания. На рис. 3 представле-ны графики N+(t), N-(t), демонстрировавшиеся автором на конференции
“Эволюция биосферы” в 1998 г. Из списка родов, использованных для пос-троения кривых, исключены археоциаты (то же на рис. 5-8, 10), что будет пояснено ниже. Прежде всего на рис. 3 обращает на себя внимание значи-тельное сходство графиков для семейств и родов. Оно проявляется в нали-чии протяженных почти прямолинейных участков, занимающих сходное положение. Так, почти прямые характеризуют появление как семейств, так и родов с начала аренига до конца турне или вымирание в юре и раннем мелу и в других случаях, хорошо видных на графиках. Для таких участков достаточно точно вычисляется усредненная (т. е. фоновая) скорость появ-ления и вымирания (разность ординат крайних точек/временной интер-
<![if !vml]>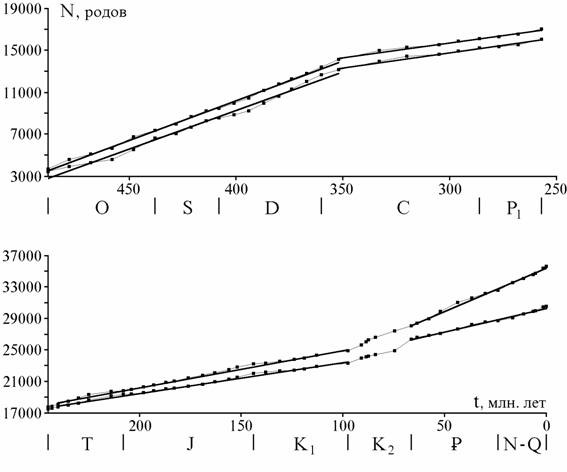 <![endif]>
<![endif]>
Рис. 4. Число появившихся (верхняя линия) и вымерших (нижняя линия) родов с линейной аппроксимацией на интервалах арениг - турне, визе - ленерд, анизий - альб, кайнозой
вал). Хорошее соответствие наблюдается в большинстве случаев и в более мелких особенностях кривых.
Во-вторых, при сравнении кривых N+(t), N-(t) для таксонов одного ранга обнаруживается близкое совпадение таких в целом почти прямоли-нейных участков (поздний палеозой и др.) или участков сходной общей конфигурации (кембрий - тремадок). Примерное сохранение разнообразия на одном уровне длительное время (в среднем и позднем палеозое) или его общее возрастание с той или иной скоростью (в раннем палеозое и мезо-кайнозое; рис. 1) определяется в результате изменениями в соотношении фоновых скоростей появления и вымирания при переходе от одного харак-терного участка кривых N+(t), N-(t) к другому. На всех четырех кривых можно грубо выделить следующие более или менее прямолинейные участ-ки: франкон - тремадок (?), арениг - турне, визе - ленерд, анизий - альб, да-ний - ныне (рис. 4). Три последние разделены поднепермско-раннетриасо-вым и позднемеловым переходными интервалами.
Таблица 1
Фоновые скорости появления и вымирания в основных частях фанерозоя
Семейства Роды
Появление Вымирание Появление Вымирание
Франкон-тремадок 4.3(0.6) 2.9(0.6) 33.1(0.5) 30.0(0.6)
Арениг-турне 8.9(1.3) 6.7(1.4) 76.6(1.2) 71.4(1.3)
Визе-ленерд 3.0(0.5) 2.9(0.6) 28.1(0.4) 28.1(0.5)
Анизий-альб 5.1(0.8) 2.6(0.6) 47.6(0.8) 38.9(0.7)
Даний-ныне 7.2(1.1) 2.0(0.4) 114.9(1.8) 62.0(1.2)
Средняя скорость 6.6 4.7 62.5 53.4
Фоновые скорости появления и вымирания для указанных пяти час-тей геохронологической шкалы даны в табл. 1 (таксонов/млн. лет; для со-поставления в последней строке приведена средняя скорость от начала томмотского века доныне, а в скобках при каждой скорости - ее отноше-ние к средней).
Числа в скобках обращают на себя внимание хорошим совпадением оценок по семействам и родам в четырех первых строках. Как и в случае существенного сходства кривых разнообразия по таксонам разного ранга, здесь выявляется характеристика, не зависящая (или слабо зависящая) от таксономического уровня изучения. Для объяснения ее происхождения достаточно предположить, что в среднем после появления (или вымира-ния) некоторого характерного числа родов появляется (вымирает) одно се-мейство. В кайнозое эта закономерность нарушается.
Существенные различия фоновых скоростей на разных интервалах фанерозоя должны быть связаны с общей организацией древних биот и за-служивают специального палеобиологического анализа, как и процессы, происходившие при переходе от одного характерного участка кривых к другому.
Локальные скорости. В отличие от средних (фоновых) скоростей на больших временных интервалах, оценка скоростей для отдельных веков затруднительна из-за недостаточной точности их продолжительностей. Это препятствует изучению так называемых “массовых вымираний”, кото-рые всегда определяются соотношениями локальных скоростей, представ-ляя собой явление второго порядка, накладывающееся на общую тенден-цию развития. Тем не менее, обсуждение некоторых аспектов вымирания оказывается возможным. В табл. 2 показаны скорости появления и выми-рания таксонов от силура до начала карбона (в скобках при скоростях - их отношение к фоновой скорости для аренига-турне).
Прежде всего следует отметить хорошее соответствие отношений к фоновой скорости: среди 22 пар чисел имеется 6 совпадений, по 5 пар от-личаются на 0.1 и 0.2, в трех случаях на 0.3, в остальных трех расхождения более существенны. В среднем разности в сравниваемых парах чисел
Скорости появления и вымирания родов в силуре - турне
Семейства Роды
Появление Вымирание Появление Вымирание
Лландовери 7.4(0.8) 3.7(0.6) 61.9(0.8) 40.2(0.6)
Венлок 13.4(1.5) 8.1(1.2) 105.2(1.4) 85.9(1.2)
Лудлов+пржидол 8.1(0.9) 11.0(1.6) 60.2(0.8) 70.6(1.0)
Жедин 6.4(0.7) 4.4(0.7) 65.5(0.9) 50.5(0.7)
Зиген 5.7(0.6) 3.3(0.5) 70.9(0.9) 45.2(0.6)
Эмс 10.9(1.2) 6.3(0.9) 108.0(1.4) 103.3(1.4)
Эйфель 7.9(0.9) 7.7(1.2) 86.7(1.1) 98.3(1.4)
Живет 8.5(1.0) 12.5(1.9) 79.8(1.0) 117.3(1.6)
Фран 6.3(0.7) 11.9(1.8) 69.7(0.9) 105.0(1.5)
Фамен 9.3(1.0) 13.4(2.0) 85.3(1.1) 88.2(1.2)
Турне 11.1(1.2) 4.9(0.7) 93.2(1.2) 56.1(0.8)
втрое больше, чем для фоновых скоростей (без кайнозоя), что вполне ес-тественно, так как здесь рассматриваются небольшие временные интерва-лы, в связи с этим не очень большое число таксонов в каждом из них и ста-тистическая представительность материала ниже. Три случая значитель-ных различий относятся к скоростям вымирания в конце силура, эмсе и фамене. В конце силура и девона интенсивность вымирания семейств по сравнению с фоновой выше, чем родов. Вероятно, здесь сказывается пре-дыдущее вымирание представителей тех же семейств, в результате которо-го к концу периода они включают меньше родов и возрастает роль моно-типных семейств. В эмсе, когда еще продолжался, постепенно замедляясь, раннедевонский рост разнообразия, в отличие от обоих терминальных вы-мираний, более интенсивным оказывается вымирание родов, а не се-мейств. В числах сравнение следующее: в эмсе к вымиранию семейства приводит вымирание в среднем 16.4 родов, в конце силура - 6.4, в фамене - 6.6 родов. Этот же эффект может сказываться при сравнении пермского и маастрихтского вымираний, в конце юры и позднем эоцене.
Массовые вымирания. При анализе собственно скоростей вымира-ния родов максимальными оказываются скорости не в конце силура или во фране и фамене, а в венлоке и живете. Скорость вымирания в лудлове и пржидоле вместе практически равна фоновой и снижается по сравнению с венлоком на 15 единиц. Минимум на кривой разнообразия у границы си-лура и девона появляется вследствие значительного, на 45 единиц, сниже-ния скорости появления родов в конце периода. По семействам, хотя в конце силура скорость вымирания возрастает на 3 единицы, основную роль в появлении минимума играет снижение скорости появления на 5 единиц. В девоне не сильно различающиеся высокие скорости вымирания родов свойственны длительному времени с эмса до конца франа. Умень-шение числа родов в это время определяется постепенным снижением скорости их появления со 108 до 70. По семействам картина в девоне не-сколько иная, но это не имеет большого значения, как и приведенные здесь все без исключения силурийско-девонские скорости вымирания и появления, слишком ненадежные, чтобы ими можно было реально пользо-ваться.
Силурийские и девонские скорости иллюстрируют в данном случае
не столько фактическую картину процесса, сколько неполную адекват-ность традиционного языка его описания. “Массовые вымирания” не обя-зательно связаны со сколько-нибудь экстраординарным вымиранием, а вы-сокие скорости вымирания могут не сочетаться с “массовым вымирани-ем”. Так, в коньяке скорость вымирания - 12 семейств и 106 родов/млн. лет (для сравнения: в кампане - 3 и 53, в маастрихте - 15 и 182). Это, конечно, может быть артефактом, зависящим от неверного определения продолжи-тельности коньяка, но очевидно “коньякское массовое вымирание” не об-суждается, поскольку здесь нет снижения разнообразия. Действительно, в коньяке скорость появления больше скорости вымирания - 23 семейства и 256 родов/млн. лет (в кампане - 5 и 81, маастрихте - 6 и 80).
Минимумы разнообразия. В истории палеонтологии и стратигра-фии “массовое вымирание” естественно ассоциируется с наибольшим раз-личием фаун до и после него, что восходит к традициям катастрофизма XIX века. Это же различие определяет положение соответствующей стра-тиграфической границы. Выраженное обособление двух последователь-ных фаун возникает, если их смене соответствует минимум разнообразия, то есть наименьшее число общих таксонов. Некоторый дискуссионный стратон, во время формирования которого разнообразие изменялось, будет в конце концов классифицирован в соответствии с тем, на какой из его границ число таксонов больше, так как это и есть таксоны, общие с фау-ной смежного века. Поэтому хронологические интервалы до минимума разнообразия объединяются в составе одной крупной геохронологической единицы, а после минимума - в составе другой. Неудивительно, что клас-сики стратиграфии проводили основные границы по минимумам разнооб-разия - такова природа самой процедуры стратиграфической классифика-ции. История переноса датского яруса из мела в палеоген хорошо иллюст-рирует процесс фиксации границы, обеспечивающий соответствие ее ми-нимуму разнообразия и преодолевающий несоответствие, которое возник-ло при фиксации по приоритету (точность соответствия зависит от формы кривой разнообразия в окрестностях минимума). Однако, высокая ско-рость вымирания перед минимумом не является условием его появления. До минимума скорость появления должна быть меньше скорости вымира-ния, а после него наоборот, но никаких других ограничений условие ми-нимума не содержит. Поэтому отождествление минимума разнообразия и “массового вымирания”, подразумевающего высокие или даже катастро-фические скорости вымирания, может оказаться некорректным.
Граница триаса и юры. В дополнение к приведенным выше приме- рам по силуру и девону можно проиллюстрировать эту ситуацию материа-лом по позднему триасу и началу юры (табл. 3), так как “массовое вымира-ние на границе триаса и юры” относится к числу хрестоматийных (в скоб-ках приведено отношение к фоновой скорости для анизия-альба).
Таблица 3
Скорости появления и вымирания родов в карнии-геттанге
Семейства Роды
Появление Вымирание Появление Вымирание
Карний 6.4(1.2) 7.4(2.8) 86.4(1.8) 82.7 (2.1)
Норий 2.5(0.5) 3.3(1.3) 38.0(0.8) 45.0 (1.2)
Рэт 3.2(0.6) 6.7(2.6) 17.4(0.4) 37.4 (1.0)
Геттанг 9.5(1.9) 1.0(0.4) 37.7(0.8) 13.2 (0.3)
Как и в конце силура, скорость вымирания родов в рэте соответству-ет фоновой, снижаясь по сравнению с карнием более чем в 2 раза. Посте-пенное уменьшение числа родов в позднем триасе определяется снижени-ем скорости их появления в 4,5 раза. Повышение скорости вымирания се-мейств в рэте (по сравнению с норием, но не карнием) связано с тем же эффектом, что и в силуре и девоне. Хотя к численным оценкам скоростей можно относиться скептически, общий вывод не вызывает сомнений: гра-ница триаса и юры соответствует минимуму разнообразия, при этом имею-щийся фактический материал не дает повода обнаружить здесь массовое вымирание. Привычное описание событий на этой границе как массового вымирания вводит в заблуждение, и от него следует отказаться.
Другие границы. Аналогичная картина изменений скоростей поя-вления и вымирания родов свойственна снижению разнообразия также в серпуховском веке и в конце юры. В ашгилле и позднем эоцене происхо-дит некоторое повышение скоростей вымирания родов, сопровождающее-ся резким снижением скорости появления.
График без временной шкалы. Как видно из предыдущего обсуж-дения, полная картина складывается при совместном анализе скоростей вымирания и появления, но труднопреодолимым препятствием при этом оказывается отсутствие надежной детальной временной шкалы (Рублев, 1996). Поэтому желательно наиболее полно использовать собственно пале-онтолого-стратиграфические данные вне зависимости от продолжитель-ностей веков. Такую возможность предоставляют графики без временной оси, в первую очередь N(N+). Наклон графика между смежными точками зависит только от отношения числа вымираний и появлений:
tga =1 - DN-/DN+.
<![if !vml]> <![endif]>
<![endif]>
Рис. 5. Изменения числа родов морских животных в координатах (N+, N)
На рис. 5 показана кривая N(N+) для родов от венда доныне (соответ-ствующую кривую для семейств см. Дмитриев, 1989). В результате исклю-чения данных по археоциатам на кривой исчез раннекембрийский макси-мум, хорошо выраженный на рис. 1 и полностью определяющийся этой группой. Исходный список содержит датировки, в большинстве случаев детализированные до подъярусов. Это не всегда удается использовать, так как в некоторых временных интервалах детализированных датировок меньше, чем неуточненных; тогда интервал рассматривался в целом (тер-минальный венд, немакит-далдынский век, пржидол, жедин, зиген, эй-фель, касимовский, гжельский, ассельский, сакмарский, джульфинский и дорашамский века, рэт, геттанг, аален, берриас, валанжин, баррем, турон, коньяк, сантон, даний, палеоцен, поздний эоцен, ранний и поздний олиго-цен, ранний, средний и поздний миоцен, плиоцен, плейстоцен). Несколько веков делятся на три части (верхи среднего кембрия, карадок, ашгилл, лландовери, фран, фамен, ленерд, анизий, норий, бат, келловей, оксфорд, альб и сеноман), остальные - на две части. Другие особенности хронологи-ческой шкалы следующие: ранний кембрий после атдабана двучленный, тойонский век отнесен к верхней части этого интервала; средний кембрий трехчленный без наименования веков; шкала перми включает ассельский, сакмарский, ленердский, гваделупский, джульфинский и дорашамский ве-ка. Всего рассмотрено 148 временных интервалов, наименьшее число уч-тенных в каком-либо из них родов - 32 (роды, вымершие в нижней части начала среднего кембрия) и 33 (появившиеся во второй половине инда). Среднее число появлений в одном временном интервале - 240, вымираний - 206 ( для сравнения то же для семейств по векам: 48 и 35).
Кривая N(N+) в целом проще и глаже кривой N(t) и, поскольку не за-висит от шкалы абсолютного времени, может быть детальнее, вплоть до зон и подзон. Детализация хронологической шкалы в данном случае поз-воляет убедиться в отсутствии, в большинстве случаев, каких-либо су-щественных событий внутри веков: дополнительные точки, как правило, не меняют общей картины. Лишь в фамене выявляется небольшой макси-мум, характеризующий развитие и быстрое исчезновение собственной фа-менской фауны. В остальных случаях удается несколько уточнить хроно-логию основных процессов. Снижение разнообразия в конце ордовика ре-гистрируется в течение среднего и позднего ашгилла; в конце силура - весь лудлов и пржидол; девонское - весь средний девон и франский век, затем в конце фамена; раннекаменноугольное - весь серпуховский век; позднека-менноугольное - только в гжельском веке; пермское - с позднего ленерда до конца периода и далее в раннеиндское время; триасовое - в позднем но-рии и рэте; позднеюрское - в течение всего титона; позднемеловое - почти полностью в позднем маастрихте, хотя незначительное снижение фиксиру-ется и в раннем маастрихте; очень слабое снижение числа родов наблюда-ется в позднем эоцене, дробнее не подразделенном. Если принять поддер-живаемую многими стратиграфами точку зрения о несамостоятельности рэта (Шевырев, 1990), минимум разнообразия на границе триаса и юры окажется единственным из крупных общепризнанных событий, для кото-рого снижение разнообразия локализуется при дроблении шкалы до подъ-ярусов в пределах одного временного интервала (в позднем нории), но при учете деления его на две зоны картина постепенного уменьшения числа родов восстановится. К сожалению, обсуждение скоростей появления и вымирания, которое было проведено выше по векам, для более детальной хронологической шкалы невозможно.
Обращает на себя внимание простота кривой N(N+) (рис. 5) с протя-женным, близким к горизонтальному участком в кембрии, плавной ордо-викской дугой, сходными циклами развития в силуре и девоне (до конца франского века) и т. д., что наглядно демонстрирует преимущества этого метода.
Эффект Сигнора-Липпса. Когда анализируются эффекты, которые могут возникать в случае катастрофических вымираний, указывают на сглаживание кривой разнообразия перед вымиранием вследствие неполно-ты имеющегося фактического материала: последние находки представите-лей некоторых таксонов, вымерших вследствие катастрофы, будут сдела-ны в более древних отложениях (Signor, Lipps, 1982). Так можно объяс-нять, например, уменьшение разнообразия в раннем маастрихте или сред-нем ашгилле, но вряд ли приемлемо распространение этого на все миниму-мы разнообразия. Катастрофические вымирания трудно предполагать для тех минимумов, которые не показывают максимальных скоростей вымира-ния в непосредственной связи с гипотетической катастрофой.
Независимость скоростей от разнообразия. Возвращаясь к графи-кам N+(t), N-(t) (рис. 3, 4), следует обсудить их общую форму. Графики не показывают какой-либо единой тенденции в изменениях скоростей на всем протяжении фанерозоя, то есть нет сколько-нибудь отчетливо выраженной зависимости скоростей от разнообразия (такой же вывод был сделан А. Гофманом (Hoffman, 1985) по семействам в результате прямого изучения корреляции скоростей и числа семейств<![if !supportFootnotes]>[1]<![endif]>). Так, в конце альба ро-дов установлено в 4.7 раза больше, чем в начале анизия, но раннемеловые скорости не больше ранне-среднетриасовых, а несколько меньше. Ско-рость вымирания родов на протяжении значительной части кембрия близ-ка к средней для всего фанерозоя, как и в кайнозое в целом, хотя мгновен-ные числа родов в кембрии и кайнозое различаются многократно.
Удельные скорости (рис. 6). Отсутствие отчетливо выраженной за-висимости скоростей вымирания и появления таксонов от разнообразия указывает на необходимость внимательного отношения при эмпирическом изучении материала к такому показателю, как удельная скорость появле-ния или вымирания. Удельные скорости должны уменьшаться при росте
<![if !vml]> <![endif]>
<![endif]>
Рис. 6. Зависимость скоростей появления и вымирания родов морских живот-ных от разнообразия (левые диаграммы относятся к появлению, правые к вымиранию): а, б - диаграммы удельная скорость - среднее мгновенное разнообразие; в, г - то же в логарифмических координатах; д, е - гистограммы отклонений логарифма удельных скоростей от его значения, вычисленного в соответствии с уравнением (5). Обозначе-ния: Nср - полусумма числа родов на границах века, t - продолжительность века, ln* - вычисленное значение логарифма, n - число попаданий разности в классовый проме-жуток
разнообразия, что действительно указывалось для вымирания на уровне семейств (Flessa, Jablonski, 1985) и может быть легко показано для родов.
На рис. 6 показана зависимость удельных скоростей появления (рис. 6, а) и вымирания (рис. 6, б) родов от среднего мгновенного числа родов для веков фанерозоя. Зависимость в целом близка к гиперболической, она линеаризуется при переходе к логарифмическим координатам (рис. 6, в, г). Гистограммы (рис. 6, д, е) показывают распределение отклонений от ли-нии регрессии логарифмов, подчеркивая некоторую нечеткость связи удельных скоростей и разнообразия, видимую также и на скаттер-диаграм-мах (коэффициент корреляции логарифмов -0.74 для появлений и -0.77 для вымираний). Уравнение регрессии:
Q=bNсрg, (5)
где Nср - полусумма числа родов на границах века, Q - удельная скорость (появления или вымирания), b, g - параметры, g < 0. Важным является па-раметр g равный -0.66 для появления родов и -0.82 для вымирания. Незави-симости скоростей от разнообразия соответствует g = -1. Для фанерозоя в целом g ¹ -1, но здесь может сказываться слишком обобщенный подход, игнорирующий неоднородность диверсификации в фанерозое, поэтому желательно рассмотреть процесс более детально.
В данном случае это оказывается удобно сделать, используя непос-редственно зависимости N+(t), N-(t) (рис. 4). Для значительных временных интервалов они удовлетворительно аппроксимируются отрезками прямых, то есть
N+(t) = a1 + b1t,
N-(t) = a2 + b2t, (6)
откуда
Q = b/N. (7)
По крайней мере для трех из указанных интервалов (кроме позднего пале-озоя) имеются достаточные основания считать скорости появления и вы-мирания родов в целом действительно независимыми от разнообразия, а удельные скорости в среднем обратно пропорциональными разнообразию. В позднем палеозое изменения числа родов незначительны, и оценка их связи с удельными скоростями затруднена.
Колебания скоростей. Линейность зависимостей N+(t), N-(t) должна иметь своим следствием линейность изменений числа родов N(t) (с разны-ми скоростями на соответствующих интервалах), но такое представление изменений разнообразия сильно огрубляет наблюдаемую картину. Погре-шность аппроксимации (6) достигает почти 500 родов в эмсе, а в среднем составляет около 100 родов. Этим можно пренебречь при оценке уравне-ний регрессии (6), так как погрешность относится к числу порядка 104, но разнообразие N(t) имеет порядок 102 - 103, а его минимумы в этом случае игнорируются. Наибольшее расхождение относится к позднему эоцену (485 родов) при среднем около 120 родов, а относительная ошибка дости-гает 25% (геттанг) и даже 46% (карний), средняя относительная ошибка 10%. Отклонения кривой разнообразия от прямой связаны с изменениями наблюдаемых скоростей появления и вымирания родов. Скорости не пос-тоянны, а колеблются около определенных средних значений. Изучая самые общие тенденции диверсификации на продолжительных интервалах геологического времени (эры или их крупные части, 108 лет), изменения скоростей можно игнорировать, но при анализе более кратковременных явлений, в частности минимумов и максимумов разнообразия с временами порядка 107 лет (эпохи), они должны учитываться.
Постоянные скорости. Модель с постоянными скоростями можно
рассматривать как идеализацию, пригодную для обобщенного описания на больших временах. Такой процесс предполагает наличие саморегуляции, поддерживающей обратную пропорциональность разнообразия и удель-ных скоростей. Вероятности появления и вымирания родов, связанные с удельными скоростями, не постоянны, как при экспоненциальном росте (модель с постоянными вероятностями см. Raup et al., 1973), а уменьшают-ся при увеличении разнообразия. Постоянная вероятность может быть приписана только всей биоте, которая контролирует процессы изменений в отдельных таксонах. Образно говоря, решение измениться или вымереть принимает не сам таксон, а за него решает биота. В этом смысле эволюци-онируют не таксоны, а биота в целом. Поскольку речь идет о единичном объекте, то диверсификация биоты является дифференцировкой, управляе-мой самой биотой. Последняя тем самым оказывается организмичным объектом (см. Чайковский, 1990; Красилов, 1992; Sole et al., 1997; Kirchner, Weil, 1998).
Объяснение постоянных скоростей в самой общей форме можно ис-кать, исходя из представлений о биоценотической регуляции эволюции, которые включают среди прочего и регулирование скоростей (Жерихин, 1987, Каландадзе, Раутиан, 1993). Идеи, лежащие в основе этой концеп-ции, связаны с представлениями о стабильности современных ненарушен-ных экосистем, то есть с временами порядка103, редко 104 лет (выше пока-зано, как изменение масштаба времен на порядок качественно меняет кар-тину процесса). Приведенные здесь результаты дают собственно палеонто-логическое подтверждение того, что регулирование скоростей в геологи-ческом прошлом действительно могло происходить и его возможные био-логические механизмы нуждаются в выяснении. Необходимо специальное более детальное исследование материала, в частности по отдельным круп-ным группам организмов.
Вероятность вымирания. Как будет проиллюстрировано далее, об-щее увеличение числа таксонов сопровождается увеличением среднего чи-сла подчиненных таксонов, в частности родов в составе семейств. Это мо-жет обусловить значительную часть снижения вероятности вымирания се-мейств в фанерозое (Flessa, Jablonski, 1985). Такой же механизм для видов может сказываться на вероятностях вымирания родов. При уменьшении среднего числа видов в составе надвидовых таксонов вероятности их вы-мирания могут увеличиваться без изменения вероятностей вымирания видов. Это указывает на необходимость дальнейшей детализации исследо-ваний до уровня видов.
Сказанное выше верно в предположении независимого вымирания видов. В этом случае снижение удельной скорости вымирания в значи-тельной части может объясняться без регулирования, выявление которого тем самым затрудняется. Если вымирание таксонов не независимо и про-исходит вследствие конкуренции между ними, то скорость вымирания должна закономерно возрастать с ростом разнообразия (пропорционально числу биотических связей между представителями разных таксонов), а удельная скорость не будет уменьшаться. Поскольку наблюдается обрат-ная картина, следует заключить, что конкуренция не является определяю-щей для скоростей вымирания. Регулирующая роль биотических отноше-ний в таком случае может состоять в ограничении конкуренции при разде-лении экологических ниш в результате эффективного действия принципа Гаузе, что может обусловить общую картину квазинезависимого вымира-ния таксонов.
Скорость появления. В отношении скорости появления новых так-сонов положение противоположное. Независимая эволюция видов будет иметь своим следствием увеличение скорости появления родов при воз-растании разнообразия, еще большее ускорение должно наблюдаться в случае зависимости появлений от конкурентных отношений. Независи-мость скорости от разнообразия указывает на подавление этих тенденций. Простейший обсуждаемый в этой связи механизм связан с ограничениями возможного дробления пространства ресурсов на экологические ниши. Тогда в течение продолжительных интервалов геологического времени в среднем сохранялся определенный баланс, с одной стороны, между ста-бильным разделением экологических ниш, ограничивающим конкурен-цию, с другой, - полнотой и дробностью использования пространства ре-сурсов, ограничивающей возможности формирования новых экологичес-ких ниш.
Экологический вакуум. Появления и вымирания таксонов в модели с постоянными скоростями всегда затруднены вне зависимости от наблю-даемого числа таксонов. В норме они происходят как случайные события с постоянной вероятностью для биоты в целом (стационарный пуассонов-ский процесс), то есть описание диверсификации на основе постоянных скоростей плохо согласуется с представлениями о бурном росте разнооб-разия после массовых вымираний (Алексеев, 1989). При переходе к удель-ным скоростям и связанным с ними вероятностям (характеристика не био-ты, а таксонов, в данном случае родов) их зависимость от разнообразия от-четливо выявляется: вероятности выше при низком разнообразии. Если изолированно рассматривать удельные скорости появления родов, свобод-ную диверсификацию в условиях “экологического вакуума” можно обнаружить, то есть в случае постоянных скоростей это понятие зависит от того, рассматривается ли процесс применительно ко всей изучаемой со-вокупности групп или к отдельному усредненному таксону.
Стабильность биоты. Обратная зависимость удельных скоростей от разнообразия определяет закономерное линейное увеличение среднего времени существования таксонов и также линейно замедляющееся обнов-ление фауны при увеличении разнообразия, то есть возрастание стабиль-ности биоты.
Локальные скорости. Реальный процесс не противопоставляется модели с постоянными скоростями, а рассматривается в рамках ее даль-нейшего развития и детализации. Поскольку аппроксимация (6) не дает возможности удовлетворительно описывать изменения разнообразия N(t) в течение веков и эпох, необходимо рассмотреть локальное поведение ско-ростей появления и вымирания. Это связано с упоминавшимися выше трудностями определения скоростей, но можно надеяться на получение результатов общего характера, мало зависящих от конкретных неточнос-тей.
Прежде всего необходимо выяснить, в какой мере отклонения ско-ростей от их средних фоновых значений могут быть обусловлены случай-ными колебаниями и погрешностями исходных данных. Появления и вы-мирания родов в модели с постоянными скоростями можно сравнить с пу-ассоновским процессом с параметром, равным фоновой скорости (в гаус-совском приближении, так как появления и вымирания родов в веке изме-ряются сотнями). Нормированное отклонение числа появлений и вымира-ний родов от их числа, предсказываемого уравнениями (6), по 17 лучше всего датированным интервалам с сеномана до ныне (без маастрихта, все-го 34 сравнения) в 26 случаях (76%) оказывается больше 2s, в 24 случаях (71 %) больше 3s и достигает почти 14s (появление родов в коньяке и вы-мирание в плиоцене), то есть изменения наблюдаемых скоростей намного больше допускаемых в стационарном пуассоновском процессе.
Влияние временной шкалы. Приведенные оценки сильно зависят от погрешностей данных как стратиграфо-палеонтологических, так и изо-топно-геохронологических. Вклад последних можно оценить, пренебрегая пока-что палеонтолого-стратиграфическими погрешностями, которые бу-дут рассмотрены ниже в специальном разделе. Если скорости появления и вымирания постоянны, относительная ошибка r продолжительности ин-тервала, достаточная для появления нормированного отклонения u, будет
r = u/(bt)1/2,
где t - принятая продолжительность интервала, b - фоновая скорость. Для позднего мела - кайнозоя (без маастрихта) определяемая таким способом средняя относительная ошибка продолжительностей 31%. Максимальная относительная ошибка - 135% , относится к появлению родов в коньяке, там же для вымираний - 60%. Продолжительность коньяка в использован-ной здесь шкале Американского геологического общества скорее всего преуменьшена, в большинстве же случаев оценки r соответствуют обыч-ным представлениям о точности изотопно-геохронологической шкалы позднего мела - кайнозоя (в палеозое разные изотопно-геохронологичес-кие шкалы нередко показывают двух-трехкратно различающиеся продол-жительности веков).
Влияние стратиграфической корреляции. Дополнительное обсто-ятельство, затрудняющее непосредственное сопоставление оценок по па-леонтолого-стратиграфическим данным с изотопно-геохронологической шкалой, связано с природой тех и других оценок. Продолжительности ве-ков в изотопно-геохронологической шкале определяются по датировкам границ, которые вычисляются на основе условия об их наилучшем соот-ветствии всем пригодным аналитическим определениям возраста из уве-ренно датированных отложений в окрестностях границы. Точное положе-ние изохронной поверхности, имеющей полученную таким способом дати-ровку, в большинстве конкретных разрезов не может быть указано, а на практике оно принимается таким, какое дано в использованной схеме стратиграфической корреляции. То же относится к оценкам по глобаль-ным палеонтоло-гостратиграфическим данным. Если корреляционные схе-мы различны, изохронные уровни заведомо не совпадают и возникнет до-полнительное несоответствие оценок.
Изменения скоростей значимы. Полностью отнести наблюдаемые отклонения скоростей появления и вымирания от постоянных на счет по-грешностей временной шкалы не удается. В этом случае относительная ошибка должна быть примерно одинакова при оценке ее и для появлений и для вымираний. Таких интервалов выявлено всего четыре: турон (32 и 35%), сантон (7 и 4%), даний (17 и 16%) и поздний эоцен (30 и 31%), ос-тальные интервалы (7 из 17) такого совпадения не обнаруживают, а часто и знак погрешности отличается (остальные 6 интервалов). В случае совпа-дения знака можно выделить ту часть большего из отклонений, которая может быть не связана с погрешностями временной шкалы, если меньшее отклонение отнести полностью на счет временной погрешности. Сделан-ные таким способом оценки несколько уменьшают нормированное откло-нение остатка, но он все равно слишком часто оказывается выходящим за пределы допускаемого для стационарного процесса. Таким образом, о су-щественных изменениях скоростей свидетельствуют: во-первых, слишком большие нормированные отклонения, не устраняемые в предположении об их хотя бы частичном возникновении в результате погрешностей времен-ной шкалы; во-вторых, разный знак погрешности для появлений и выми-раний в некоторых интервалах; в третьих, наличие минимумов на кривой разнообразия.
<![if !vml]>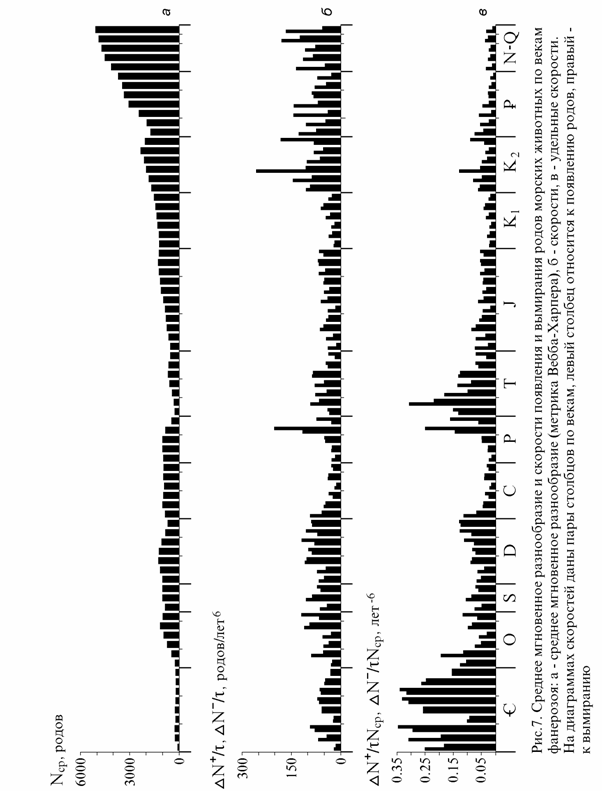 <![endif]>
<![endif]>
По крайней мере частично изменения наблюдаемых скоростей могут быть связаны с погрешностями палеонтолого-стратиграфических данных. Последние в данном разделе не учитываются, а в отношении скоростей ниже приведены результаты дальнейшего анализа, необходимость которо-го следует из предположения об их неслучайных изменениях.
Обзор локальных скоростей. Рис. 7 дает представление об измене-ниях скоростей и удельных скоростей по векам фанерозоя. Обращает на
себя внимание волнообразное, в большинстве случаев согласованное уве-личение или уменьшение скоростей и удельных скоростей (рис. 7, б, в) как появления, так и вымирания с постепенными, в течении обычно несколь-ких веков, изменениями в их соотношении (см. также рис. 5). Общий ха-рактер этой картины не изменится при уточнении продолжительностей ве-ков, которые связаны общей продолжительностью более длительных вре-менных интервалов и могут только перераспределяться в ее пределах. Со-отношение скоростей в отдельных веках не зависит от их продолжитель-ностей. Уточнение последних может изменить лишь относительную высо-ту пар столбцов в разных веках (рис.7, б, в) из числа составляющих наибо-лее надежно датированные более длительные интервалы и несколько смес-тить максимумы и минимумы. Так как изменения скоростей и удельных скоростей появления и вымирания координированы, представляет интерес их разность, определяющая собственно изменения разнообразия.
Затухающие колебания ? На рис. 8 показана удельная скорость из-менений разнообразия по векам. Видны четыре однотипные серии нерегу-лярных затухающих колебаний: кембрийская, ордовикско-раннепермская, позднепермско-раннемеловая и позднемеловая-кайнозойская. Кембрийс-кая своеобразна, а последняя не завершена, тогда как две другие вероятно отображают полные циклы диверсификации. Они отличаются затуханием колебаний около 0 в позднем палеозое (постоянное разнообразие) и в об-ласти небольших положительных значений в раннем мелу (медленно рас-тущее разнообразие) на фоне юрско-раннемелового общего снижения удельной скорости диверсификации. Ненулевая величина ее в раннем мелу может быть связана с участием в составе фауны мезозоя и кайнозоя все большего числа (по мере приближения к современности) существующих ныне родов (см. ниже раздел о мерах разнообразия). Хотя кривая может в дальнейшем подвергаться значительным уточнениям (в частности, поздне-меловой максимум снизится ?) , ее общий характер вряд ли изменится принципиально.
Как известно, многим системам с регулированием свойственны де-терминированные автоколебания различного типа, обусловленные времен- ными задержками в контурах обратной связи, в том числе затухающие, пе-риодические или квазистохастические. Кривая на рис. 8 вполне может ото-бражать процесс такого типа с возможным существенным участием сто-хастической составляющей. В этой связи каузальный анализ изменений разнообразия может значительно усложниться.
Влияние внешних факторов. Рис. 8 косвенно указывает на возмож-ность единообразного количественного описания глобального процесса диверсификации, включая эпизоды снижения разнообразия, на основе мо-делей, в которых влияния абиотических факторов не требуется (хотя оно не вызывает сомнений на региональном уровне - неоген Паратетиса и др.). Удачное построение таких моделей (с биологически оправданным истол-кованием смысла и величины параметров) означало бы невозможность вы-
<![if !vml]> <![endif]>
<![endif]>
Рис. 8. Удельная скорость изменения числа родов морских животных в фанеро-зое
яснить реальное значение абиотических изменений путем анализа только суммарных фаунистических данных. Для этого необходима комплексная реконструкция конкретных процессов и событий, что обычно и делается (хороший пример - изучение импактного события на границе маастрихта и дания), но исходные позиции многих таких исследований основаны на априорной убежденности в необходимости внешнего влияния: выясняется насколько тот или иной гипотетический фактор мог быть причиной гло-бального вымирания. (Реконструкция событий необходима при анализе каждого шага диверсификации вследствие постоянной смены таксономи-ческих групп).
В этой связи обращают на себя внимание ордовикско-раннепермс-кий цикл диверсификации (рис. 8). Он не связан с предваряющим сниже-нием разнообразия, то есть нет необходимости искать предшествующее крупное абиотическое событие, а последующее развитие до середины пер-ми предположительно может быть описано как детерминированные авто-колебания. На графике сходно выглядит позднемеловой-кайнозойский цикл, но его характеристики могут быть искажены: продолжительность коньяка возможно сильно занижена, а маастрихтские события имеют уни-кальные характеристики и их описание как автоколебаний может быть не-адекватным. Классические представления о вынужденных колебаниях раз-нообразия со снижением его под действием внешних причин и последую-щей мощной диверсификацией в условиях экологического пространства, свободно доступного для освоения, описывают не единственный правдо-подобный механизм изменений разнообразия. Характерно появление в последнее время гипотез о биологических процессах, связанных с общей структурой экологических ниш, в которых уменьшение разнообразия мо-жет быть слабо зависящим от абиотических событий или даже не завися-щим от них (Каландадзе, Раутиан, 1993). Здесь проиллюстрирован иной, очень простой подход к выяснению общих особенностей диверсификации, который приводит к похожим предположениям и, кроме того, открывает некоторые возможности количественного изучения. Наиболее сложны для изучения должны быть смешанные случаи, сочетающие вынужденные из-менения разнообразия с автоколебаниями.
Предположение об автоколебаниях биоразнообразия с возможно-стью его постепенного снижения в результате биотических процессов за-вершает обобщенный перечень гипотез о причинах возникновения мини-мумов разнообразия. Ранее предлагались в основном гипотезы реактивно-го снижения разнообразия в ответ на какие-либо изменения во внешней среде – постепенного (как в крупных трансгрессивно-регрессивных цик-лах), либо катастрофического (импактные события и проч.). Позже были описаны гипотетические внутрибиотные механизмы автономного возник-новения биоценотических кризисов в обшем катастрофического характера (Жерихин, 1978). Автономное постепенное снижение разнообразия приз-навалось невозможным, и последний из четырех логически возможных вариантов не обсуждался.
Режимы диверсификации. Вновь обращаясь к уравнению регрес-сии (5) (стр. 26) для удельных скоростей на больших временых интерва-лах, следует сказать, что в зависимости от значений параметров могут осуществляться весьма различные режимы диверсификации. В палеонто-логической литературе для фанерозоя в целом или его крупных частей рас-сматривались в основном два случая:
1) g1=g2=0 (индекс 1 относится к появлению, 2 - к вымиранию): экспонен-циальное увеличение разнообразия (Benton, 1995), скорости появления и вымирания ~N, удельные скорости постоянны, появления и вымирания ро-дов независимы;
2) g1=0, g2=1: ограниченное логистическое увеличение разнообразия (Cour-tillot, Gaudemer, 1996); скорость появления как в предыдущем случае, ско-рость вымирания ~N2 (или содержит слагаемое ~N2, причем более высо-кие степени N отсутствуют; этот член появляется при учете биотических отношений между представителями таксонов по аналогии с моделями за-висящего от плотности логистического роста популяций; при логистичес-ком росте слагаемое ~N2 может быть и в скорости появления), удельная скорость вымирания ~N (или линейна по N, то же для появлений).
Приведенные выше результаты не подтверждают развитие разнооб-разия морских животных по какой-либо из этих двух моделей (возможно g » 0 в кембрии). В течении значительных частей фанерозоя осуществля-лась диверсификация с g1 = g2 » -1 при колебаниях скоростей около сред-них b1, b2, удельные скорости ~N-1, основные перестройки процесса связа-ны с изменениями b1, b2.
На больших временах процесс диверсификации обнаруживает значи-тельную независимость от изменений, происходящих в течение немногих веков. Ведущие факторы, определяющие его основные особенности на временных интервалах различной характерной продолжительности, долж-ны быть разными. Рост разнообразия не может быть неограниченным, от-сутствие признаков его замедления в мезокайнозое заставляет ставить воп-рос о причинах этого явления и адекватном описании ограниченного рос-та, в том числе о его пределах и удаленности от них современной фауны.
Изменения удельных скоростей. В исследованиях изменений так-сономического разнообразия базовыми величинами являются удельные скорости появления и вымирания таксонов. Однотипная зависимость удельных скоростей от разнообразия определяет их заметную связь между собой (рис. 7, коэффициент корреляции 0.90). Если исключить этот фак-тор, изучая связь отклонений удельных скоростей от линии регрессии (7) на четырех указанных выше временных интервалах, для которых скорости появления и вымирания можно принять постоянными (60 пар значений), коэффициент корреляции 0.69. Изменения удельных скоростей обычно происходят более или менее согласованно, в том числе в окрестностях минимумов разнообразия. Отчетливо выделяются удельные скорости вы-мирания в гвадалупии и в маастрихте, намного превосходящие в этих двух случаях удельные скорости появления. Последние обычно не обнаружива-ют сколько-нибудь существенного возрастания после минимумов разнооб-разия. Так, в лландовери удельная скорость появления родов выше, чем в ашгилле, но ниже, чем в карадоке (при использовании изотопно-геохроно-логической шкалы Американского геологического общества, 1989 г). В жедине удельная скорость появления родов не выше позднесилурийской; в геттанге она меньше карнийской; в турне, башкирском веке, берриасе, раннем олигоцене ниже, чем в фамене, серпуховском веке, в конце юры и позднем эоцене соответственно. Только после пермо-триасового вымира-ния эта скорость оказывается очень высокой в оленекское время, тогда как в дании она сравнима с сеноманской и туронской. Повидимому, удельные скорости появления после минимумов разнообразия в целом укладывают-ся в общую зависимость от разнообразия и не являются необычными, то есть представления о свободной быстрой дивергенции после “массовых вымираний” не подтверждаются, хотя в этом отношении желателен более детальный анализ.
Применение к массовым вымираниям. Общая тенденция к увели-чению удельных скоростей с уменьшением разнообразия создает дополни-тельные трудности при попытках использовать удельную скорость выми-рания в качестве критерия для выявления крупных вымираний. Например, по имеющимся данным в эйфеле - фране картина удельных скоростей сле-дующая:
Появление Вымирание
Эйфель 0.071 0.081
Живет 0.075 0.110
Фран 0.084 0.127
В живете удельная скорость вымирания возрастает по сравнению с эйфельской значительно больше, чем удельная скорость появления, но дальнейшее увеличение обеих скоростей во фране сопоставимо (на 12% и 15% по сравнению с живетом) и определяется преимущественно уменьше-нием разнообразия. Несущественное трехпроцентное различие зависит от несколько большего снижения во фране абсолютной скорости появления, чем вымирания. Такие случаи затрудняют изолированное использование удельной скорости вымирания как показателя экстраординарных событий, требуя вычленения из ее общего прироста той части, которая связана с об-щей зависимостью от уменьшения разнообразия. Поскольку связь скорос-ти с разнообразием кореллятивная это, как правило, окажется затрудни-тельным. Более интересны могут быть случаи, когда удельная скорость из-меняется противоположно общей фанерозойской тенденции. Так, в карнии – рэте скорости следующие:
Появление Вымирание
Карний 0.13 0.12
Норий 0.06 0.07
Рэт 0.03 0.07
Хотя разнообразие уменьшается, возрастания удельных скоростей установить не удается, они уменьшаются, причем скорость появления с карния до рэта более чем вчетверо, вымирания - менее чем вдвое. Сходная картина наблюдается в раннем карбоне и возможно в конце силура. В кон-це юры удельная скорость вымирания не повышается, а появления снижа-ется. В эоцене удельная скорость вымирания постоянно незначительно возрастает в течение всей эпохи, тогда как появления - постоянно снижа-ется. Все сравнения скоростей и удельных скоростей в разных веках, как неоднократно указывалось выше, зависят от погрешностей изотопно-гео-хронологической шкалы и могут сильно меняться при ее уточнении. Пола-гая равновероятными уточнения продолжительностей отдельных веков как в сторону ее увеличения, так и уменьшения, следует ожидать, что качест-венные характеристики не всех, а только некоторых эпизодов могут в дальнейшем измениться на противоположные.
Высокие удельные скорости. Синхронное повышение обеих удель-ных скоростей в некоторых веках означает ускоренное обновление фау-ны. Эти века могут характеризоваться повышенной долей вымерших так-сонов без уменьшения разнообразия (плинсбах, апт, сеноман, средний ми-оцен) (Дмитриев, 1989). На кривых когортного анализа (Raup, 1978) и об-ратных лайелевых кривых эпизоды ускоренного обновления отражаются ступенеобразными изгибами. В связи с повышенной вероятностью выми-рания для таких веков должно быть характерно присутствие большего ко-личества короткоживущих таксонов, особенно в окрестностях минимумов разнообразия и в кембрии.
Диверсификация насекомых изучалась в сравнительном плане для анализа влияния, которое оказывает на кривые разнообразия использова-ние различного исходного материала. В данном случае сравнивались ре-зультаты, полученные по данным Лаборатории членистоногих ПИН и по списку К. Лабандейра (Labandeira, 1994). Различия списков очень велики. Их можно охарактеризовать следующими числами: всего семейств - 1049 (Лаб. членистоногих; из них отсутствуют в списке Лабандейры - 148) и 1263 (Лабандейра; из них отсутствуют в списке Лаб. членистоногих - 362); общих семейств для обоих списков - 901, для них датировки появления совпадают в 263 случаях (29%); в том числе вымерших общих семейств - 403, для них датировки вымирания совпадают в 170 случаях (42%).
Несмотря на столь значительные несовпадения списков, общая кон-фигурация обеих кривых разнообразия (рис. 9) оказывается весьма сход-ной. Формальная оценка сходства по коэффициенту корреляции дает 0,986 - очень высокую величину. Особый интерес представляют наиболее замет-ные различия этих двух кривых, которые указаны на рис. 9 затемненными зонами в пяти возрастных интервалах.
Два случая - в поздней юре и палеогене, демонстрируют смещение однотипных резких ступенеобразных изгибов обеих кривых на один ин-тервал. Оба ступени связаны с крупнейшими уникальными местонахожде-ниями ископаемых насекомых - Каратау в поздней юре и насекомыми бал-тийского янтаря в палеогене. Оба местонахождения не сопоставимы по объему и разнообразию коллекций с ближайшими по возрасту более древ-
<![if !vml]> <![endif]>
<![endif]>
Рис. 9. Сопоставление кривых числа семейств ископаемых насекомых: а - по К. Лабандейра (Labandeira, 1994), б - данные Лаборатории членистоногих ПИН. Тенями выделены интервалы наиболее существенных несовпадений. Объяснения в тексте
ними и более молодыми местонахождениями и содержат большое коли-чество представителей семейств, не встреченных в более древних отложе-ниях. Изученность ископаемых насекомых поздней юры и палеогена ока-зывается весьма неравномерной, что влечет за собой резкий излом кривых разнообразия в хронологических интервалах, к которым относятся эти местонахождения. Различия в положении описываемых ступеней опреде-ляются разными датировками местонахождений. Каратау в материалах Лаб. членистоногих отнесено к келловею, Лабандейра - к оксфорду; бал-тийский янтарь соответственно к позднему эоцену и раннему олигоцену.
В двух других случаях - в конце триаса и раннего мела, направлен-
ность сравниваемых кривых различна: некоторое снижение разнообразия в конце триаса по данным Лаб. членистоногих и повышение по данным Ла-бандейра; малозаметное повышение разнообразия в конце раннего мела по данным Лаб. членистоногих и его существенное снижение у Лабандейра.
В первом из них, триасовом при анализе исходного материала обна-руживается различие датировок австралийского местонахождения Ипсвич: карний у Лаб. членистоногих, рэт у Лабандейра. Поскольку в Ипсвиче за-фиксированы первые находки представителей 10 семейств, различие дати-ровок оказывается определяющим; дополнительно следует упомянуть 2 семейства из нижнеюрского местонахождения Согюты, появление кото-рых отнесено Лабандейра также к рэту в соответствии с более ранними да-тировками этого местонахождения (Беккер-Мигдисова, 1962).
Во втором случае также выявляется, что расхождение кривых зави-сит от разного понимания возраста местонахождений. В апте и альбе, если рассматривать их совместно, число семейств насекомых незначительно возрастает на обеих кривых. Различия определяются значительно более резким ростом числа семейств в апте с появлением локального максимума по данным Лабандейра. Аптский рост определяется здесь появлением 11 семейств в крупном забайкальском местонахождении Байса, которое в списках Лаб. членистоногих учитывается как поздненеокомское. Дополни-тельно к этому для альба играет роль вымирание ряда семейств, последний раз в геологической летописи встреченных в Забайкалье, в отложениях дискуссионного возраста, относимых одними исследователями к поздней юре (Пономаренко, 1990), другими - к альбу (Жерихин, 1978). Первый ва-риант принят при построении кривой по данным Лаб. членистоногих, тог-да как Лабандейра использована вторая датировка. Соответственно, ниж-няя кривая показывает снижение разнообразия в оксфорде, уменьшая тем самым число семейств, вымерших в альбе, а верхняя отражает противопо-ложную точку зрения.
Наконец, пермский участок различия двух кривых связан в основном с датировками трех крупных местонахождений: Обора в Чехии, Элмо в Канзасе и Калтан в Кузбассе. Первое ранее датировалось как ассельское, и эта датировка принята Лабандейра, тогда как в списке Лаб. членистоногих оно считается артинским, а местонахождения Элмо и Калтан отнесены не к сакмарскому ярусу, как у Лабандейра, а к артинскому первое и уфимско-му второе.
Таким образом, сопоставление результатов обработки двух незави-симо составленных списков семейств насекомых не дало возможности вы-явить существенные различия кривых, зависящие от принимаемой систе-мы, в том числе разного понимания состава семейств. Все рассмотренные случаи таких различий оказываются связаны с разными датировками круп-ных местонахождений. Неравномерная изученность ископаемых насеко-мых с непропорционально высокой ролью материала из уникальных мес-тонахождений приводит к появлению заведомых искажений общей формы кривой.
Первая интерпретация кривых разнообразия
Выше кратко охарактеризованы и проиллюстрированы некоторые практические приемы изучения изменений таксономического разнообра-зия, но принципиальные основы такого изучения неочевидны. Рассмотрен-ные выше кривые для родов, семейств и отрядов морских животных, а так-же семейств насекомых позволяют считать, что выявляемая с помощью кривых разнообразия картина его изменений в некоторых основных чер-тах слабо зависит от таксономического ранга и принимаемой системы. Это хорошо согласуется с неоднократными указаниями в палеонтологической литературе на те же свойства кривых , а также на значительную устойчи-вость их общей формы в процессе изучения (Sepkoski et al., 1981; Maxwell, Ben-ton, 1990; Sepkoski, 1993b).
Форма кривой определяется безразмерным отношением ординат, то есть число таксонов само по себе не имеет значения. Оно используется только как средство для измерения некоторой величины. Как можно пред-полагать, эта величина - морфологическое разнообразие. Какое-то соответ-ствие морфологического и таксономического разнообразий - тривиальный факт, но следует разъяснить, в каком смысле можно говорить именно об измерении первого с помощью второго и обсудить основные особенности такого измерения.
Таксону можно поставить в соответствие некоторую меру морфоло-гического разнообразия, которую удобно называть его морфологическим объемом (или просто объемом, отличая его от объема таксона как термина свободного пользования, подразумевающего чаще всего число подчинен-ных таксонов). Таксоны предполагаются различающимися по своему объе-му, следовательно вся их совокупность может описываться некоторым ста-тистическим распределением объемов и соответствующим средним объе-мом таксона. Исходя из этих общих предпосылок, можно обсудить неко-торые следствия предположения об измерении, не определяя пока-что са-мой меры.
Трудность интерпретации кривых числа таксонов как отображаю-щих изменения морфологического разнообразия определяется тем, что здесь не вводится в явной форме стандартная единица измерения; единица появляется только как средний объем таксона. При подсчете большого чи-сла таксонов становится возможным соизмерение морфологических разно-образий на разных хронологических уровнях, но оно имеет свои специфи-ческие особенности.
Таксоны во многих случаях удобно рассматривать как группы точек в многомерном пространстве признаков, это представление используется в дальнейшем изложении. Разнообразие организмов в пространстве призна-ков отображается множеством точек, которое изменяется во времени. Предполагается естественная структурированность этого множества, зада-ющая определенные рамки его разбиению по таксонам. В пределах более или менее четко очерченных таксонов систематик скорее всего стремится уменьшить дисперсию распределения объемов подчиненных таксонов. Каждый систематик работает в ограниченной части пространства призна-ков, в той или иной мере согласовывая свои решения со специалистами, занимающимися этой же частью. Это приводит к тому, что средний объем таксонов одного ранга может существенно отличаться в разных частях геохронологической шкалы даже в одной крупной группе организмов. Для разных крупных групп отсутствуют процедуры соизмерения объема таксо-нов, поэтому невозможно что-либо утверждать о соотношениях объемов. Когда строится график числа таксонов того или иного ранга, то, с точки зрения измерения морфологического разнообразия, это означает, что неяв-но принято допущение о среднем объеме таксонов этого ранга в качестве единицы для всего изучаемого возрастного интервала и для всех суммиро-ванных при составлении графика групп.
Вряд ли график числа таксонов, построенный на такой основе, не-пременно правильно отображает реальные изменения морфологического разнообразия, но можно указать условие, при соблюдении которого он бу-дет пригоден: если характерное время возможного существенного измене-ния мгновенного среднего объема таксонов много больше продолжитель-ности веков, то график верен для смежных точек кривой. Так, показанное на рис. 1, в увеличение разнообразия морских животных в 3.89 раза в нача-ле маастрихта по сравнению с началом лланвирна не следует принимать всерьез, но длительное сохранение разнообразия в палеозое и, возможно, длительный мезокайнозойский рост достаточно обоснованы, как и более частные особенности кривой - участки возрастания, убывания и стабиль-ности, главные минимумы и максимумы. Именно эти качественные хара-ктеристики представляют основной интерес, но следует убедиться в мед-ленном изменении мгновенного среднего объема таксонов.
При изучении числа таксонов невозможно определить скорость из-менения их мгновенного среднего морфологического объема (для этого нужно непосредственно использовать меру разнообразия), но косвенное суждение возможно по числу подчиненных таксонов (объему в обычном понимании). На рис. 10 показано изменение среднего числа семейств и ро-дов в составе отрядов и родов в составе семейств. Обращает на себя вни-мание присутствие на всех трех графиках тех же основных минимумов и максимумов, что и на кривых числа таксонов. При увеличении (или сниже-нии) числа таксонов как правило увеличивается (снижается) также и сред-нее число подчиненных таксонов. Кривая для числа таксонов высокого ранга, не учитывающая этого, оказывается сглаженной по сравнению с кривой для таксонов более низкого ранга. Сглаживание неоднократно вос-производилось на компьютерных моделях (Lasker, 1976). Например, отно-шение числа отрядов в конце и начале ашгилла 0.95, семейств - 0.86, родов - 0.60. В серпуховском веке число отрядов не изменяется в отличие от его увеличения в турне и визе (рис. 1), а число семейств и родов уменьшается. Прекращение (и замедление ?) роста числа таксонов высокого ранга может быть признаком снижения числа таксонов более низкого ранга. Так, палео-
<![if !vml]> <![endif]>
<![endif]>
Рис. 10. Изменения мгновенного среднего числа таксонов морских животных в составе таксонов более высокого ранга: а - родов в отряде, б - семейств в отряде, в – родов в семействе
цен-эоценовое увеличение числа родов заканчивается к позднему эоцену, что может указывать на уменьшение числа видов в позднем эоцене, тоже можно сказать о конце ранней юры и в некоторых других случаях. В тито-не наблюдается даже уменьшение числа родов при продолжающемся рос-те числа семейств и отрядов; такие случаи могут быть выявлены только при переходе к изучению следующего, более низкого уровня таксономи-ческой иерархии.
Изменения среднего числа подчиненных таксонов свидетельствуют о невысокой точности оценки изменений разнообразия с помощью числа таксонов. В течение века число родов в составе семейства меняется (по от-ношению к числу родов на его нижней границе) в среднем на 10%, то есть именно с такой средней точностью приращения числа семейств за век про-орциональны приращениям числа родов. Это неплохо для палеонтологи-ческого материала, однако в окрестностях минимумов разнообразия сред-нее число родов может меняться на 20-40%, как в сторону уменьшения слева от минимума, так и увеличения справа от него. Значительные изме-нения происходят также в раннем и среднем кембрии. Если исключить эти случаи, число родов в составе семейства меняется в течение века в сред-нем на 6%. Своеобразие кембрийской диверсификации, выявляющееся также на рис. 2, свидетельствует о необходимости специального анализа кембрийских преобразований разнообразия. В течение длительного време-ни до середины мела среднее число подчиненных таксонов меняется не-значительно, затем быстро увеличивается (кроме маастрихта и поднего эоцена). Этим объясняется несоответствие кайнозойских фоновых скорос-тей появления и вымирания семейств и родов, отмеченное выше: для окон-чательного вымирания семейства требуется все больше вымерших родов (рис. 10,в) и видов (Flessa, Jablonski, 1985). Фоновая скорость вымирания родов в кайнозое несколько выше средней для фанерозоя в целом, тогда как скорость вымирания семейств значительно ниже.
Хотя локальные приращения для таксонов разного ранга могут быть примерно пропорциональны, кривые в целом связаны нелинейно и имеют разную общую форму, что свидетельствует о возможности преимущест-венно только качественного анализа. Различие формы кривых биологичес-ки определяется историей формирования и исчезновения архетипических структур, а в терминах пространства признаков - историей общей структу-ры множества точек, отображающего мгновенные разнообразия. Так, уве-личение числа отрядов в кембрии и ордовике до 75% современного числа представленных в ископаемом состоянии отрядов в пространстве призна-ков отразится как быстрое формирование более или менее изолированных подмножеств точек.
Принципиальное ограничение метода состоит в том, что необходимо использовать оценки мгновенного разнообразия, тогда как кривые числа таксонов маскируют изменения их мгновенных средних объемов. Именно вследствие этого появляются различия кривых по таксонам разного ранга и возникает необходимость анализировать изменения среднего числа под-чиненных таксонов. Частичное решение этой проблемы при работе с так-сонами видится в увеличении разрешающей способности метода, то есть переходе на уровень видов, тогда как полное решение связано с перспек-тивой непосредственного использования мер разнообразия. Для видов ис-следование пока-что возможно только на ограниченном материале, но по мере развития быстро накапливающихся компьютерных баз данных станет более доступным.
Рис. 10 и изложенное выше его обсуждение иллюстрируют общий подход к анализу на разных уровнях детальности. Сказанное о среднем морфологическом объеме таксонов, подразумевающем максимальную де-тальность характеристики разнообразия, может быть без изменений пере-несено на уровень видов. Очевидно, имеется статистическое распределе-ние надвидовых таксонов одного ранга по числу входящих в их состав ви-дов с соответствующим средним. Как и ранее, условием пригодности кри-вой разнообразия для дальнейшего анализа будет достаточно медленное изменение мгновенного среднего числа видов, тогда мгновенное число надвидовых таксонов локально изменяется примерно так же, как число ви-дов (Дмитриев, 1989). Для имеющихся палеонтологических данных сред-нее число видов может быть подсчитано как мгновенная характеристика для границ геохронологических подразделений, так что изменения эмпи-рического среднего доступны для изучения. Аналогично можно говорить о родах в составе семейств или отрядов и семействах в составе отрядов (рис. 10). Отличие от интерпретации кривых через меру морфологического раз-нообразия заключается в том, что число подчиненных таксонов может рас-сматриваться только в рамках какого-либо конкретного варианта системы. Мера разнообразия должна быть независящей (или слабо зависящей) от принятой системы и вследствие этого характеристикой более высокого уровня общности.
Распределение таксонов по числу подчиненных таксонов хорошо из-вестно для крупных групп современных животных и растений как распре-деление Ципфа (Виллиса, Парето и др.). Оно удовлетворительно описыва-ет таксономическую структуру многих высших таксонов, поэтому в исто-рическом развитии должно быть достаточно консервативным. Распределе-ние Ципфа не центральное (мода не совпадает со средним), что обусловли-вает определенную специфичность работы с характеризуемыми им сово-купностями. На палеонтологическом материале распределение почти не изучалось, хотя представляет несомненный интерес, так как только пале-онтологические данные позволяют сопоставлять распределения для всей истории существования крупных групп или значительных временных ин-тервалов и мгновенные.
В теории ветвящихся случайных процессов мгновенные и интерваль-ные распределения связываются определенными соотношениями, справед-ливыми для некоторых размножающихся и гибнущих частиц или квази-частиц. Общие параметры участвуют также в описании распределения продолжительностей существования частиц (квазичастиц) и распределе-ния во времени точек дивергенции и филетического вымирания. Соответ-ствие (или несоответствие) эмпирического палеонтологического материа-ла теоретическим соотношениям для ветвящихся процессов представляет большой интерес в различных аспектах, в том числе в изучении природы таксонов.
Другая интепретация кривых разнообразия основана на анализе при-ращений числа таксонов в течении века как суммы приращений для от-дельных групп. На рис. 11-19 показаны приращения по основным круп-ным группам морских животных от позднего венда до современности. Ра-нее такие же диаграммы были охарактеризованы для семейств, но проил-люстрированы только выборочно (Дмитриев, 1989). Для родов, которых на порядок больше, чем семейств, удается показать на одной диаграмме от 30 (в триасе) до 50 (в девоне) групп, и общая картина более представительна (по семействам - около 15 групп). На всех приведенных здесь диаграммах масштаб одинаков, высота наименьшего столбца равна одному роду. По-ложительные приращения показаны в верхней полуплоскости, отрицатель-ные - в нижней. При построении диаграмм использованы только данные о родах, появление и вымирание которых датировано с точностью до века или его части; всего таких родов оказалось 29313.
Общая структура всей совокупности диаграмм не отличается от по-лученной ранее для семейств. При возрастании разнообразия диаграмма в целом помещается в верхней полуплоскости в результате согласованного роста разнообразия в большинстве групп. В нижней полуплоскости заня-тая диаграммой площадь незначительна, что определяется небольшим пре-обладанием вымирания в немногих группах. При убывании разнообразия диаграмма перемещается в нижнюю полуплоскость, в немногих группах может быть некоторое преобладание появления новых родов. Степень вы-раженности этой картины прямо зависит от отношения DN-/DN+, опреде-ляющего наклон графика N(N+). Увеличение DN-/DN+ соответствует пере-мещению все большей части диаграммы вниз. Если разнообразие не меня-ется (DN-/DN+ = 1), приращения для разных групп обычно невелики и хао-тически распределяются внизу и вверху. В связи с таким строением диа-грамм их смена от века к веку хорошо согласуется с ходом графика N(N+). Например, девонская дуга (рис. 5) на диаграммах для жедина - франа (рис. 13, а-е) отображается как закономерное плавное перемещение основной площади диаграммы из верхней полуплоскости в нижнюю.
Таким образом, имеется согласованное коллективное поведение всей совокупности рассмотренных групп, явно определяющееся существовани- ем факторов, регулирующих разнообразие морских животных в целом. На-ряду с обсуждавшейся выше независимостью скоростей появления и вы-мирания родов и семейств от разнообразия, это еще один показатель высо-кой интегрированности поведения всей биоты. В отличие от скоростей, в
<![if !vml]> <![endif]>
<![endif]>
Рис. 11. Приращения числа родов в основных крупных группах морских живот-ных в позднем венде и кембрии; а - поздний венд, б - немакит-далдын, в - томмот, г - атдабан, д - конец раннего кембрия, е-з - средний кембрий: е - начало, ж - середина, з - конец, и - дресбач, к - франкон, л - тремпельон. Основные группы слева направо: 1 – Protozoa, 2 - Demospongia, 3 - Calcarea, 4 - Hexactinellida, 5 - Porifera inc. cl., 6 – Regula-res, 7 - Irregulares, 8 - Trilobozoa, 9 - Cyclozoa, 10 - Scyphozoa, 11 - Hydrozoa, 12 – Antho-zoa, 13 - Coelenterata inc. sed., 14 - Chaetognatha, 15 - Annelida, 16 - Coeloscleritophora, 17 - Tommotiida, 18 - Machaeridia, 19 - прочие Problematica, 20 - Tergomya, 21 – Helcionel-loida, 22 - Rostroconchia, 23 - Orthothecimorpha, 24 - Hyolithomorpha, 25 - Bivalvia, 26 - Gastropoda, 27 - Cephalopoda, 28 - прочие Mollusca, 29 - Olenellida, 30 - Agnostida, 31 - Redlichiida, 32 - Corynexochida, 33 - Odontopleurida, 34 - Asaphida, 35 - Ptychopariida, 36 - прочие Trilobita, 37 - Ostracoda, 38 - прочие Arthropoda, 39 - Inarticulata, 40 - Articulata, 41 - Echinodermata, 42 - Pterobranchia, 43 - Graptolithina, 44 - Conodonta
<![if !vml]> <![endif]>
<![endif]>
Рис. 12. Приращения числа родов в основных крупных группах морских живот-ных в ордовике и силуре; а - тремадок, б - арениг, в - лланвирн, г - лландейло, д – кара-док, е - ашгилл, ж - лландовери, з - венлок, и - лудлов, к - пржидол. Основные группы слева направо: 1 - Protozoa, 2 - Stromatoporoidea, 3 - прочие Porifera, 4 - Tabulata, 5 – Rugosa, 6 - прочие Coelenterata, 7 - Annelida, 8 - Problematica, 9 - Hyolithomorpha, 10 – Bivalvia, 11 - Archaeogastropoda, 12 - прочие Gastropoda, 13 - Orthocerida, 14 – Actino-cerida, 15 - Endocerida, 16 - Oncocerida, 17 - прочие Cephalopoda, 18 - прочие Mollusca, 19 - Asaphida, 20 - Ptychopariida, 21 - Proetida, 22 - Phacopida, 23 - прочие Trilobita, 24 - Merostomata, 25 - Palaeocopida, 26 - прочие Ostracoda, 27 - прочие Arthropoda, 28 – Tre-postomata, 29 - Cryptostomata, 30 - прочие Bryozoa, 31 - Inarticulata, 32 - Orthida, 33 – Strophomenida, 34 - Pentamerida, 35 - прочие Articulata, 36 - Rhombifera, 37 - Diploporita, 38 - Stylophora, 39 - Crinoidea, 40 - прочие "Pelmatozoa", 41 - Edrioasteroidea, 42 - прочие "Eleutherozoa", 43 - Graptolithina, 44 - Conodonta, 45 - Cephalaspidomorphi, 46 – Pteraspidomorphi, 47 - Acanthodii
<![if !vml]> <![endif]>
<![endif]>
Рис. 13. Приращения числа родов в основных крупных группах морских живот-ных в девоне; а - жедин, б - зиген, в - эмс, г - эйфель, д - живет, е - фран, ж - фамен. Основные группы слева направо: 1 - Spumellaria, 2 - Chitinozoa, 3 - Foraminifera, 4 – Stromatoporoidea, 5 - прочие Porifera, 6 - Rugosa, 7 - Tabulata, 8 - прочие Coelenterata, 9 - Polychaeta, 10 - Machaeridia, 11 - Bivalvia, 12 - Archaeogastropoda, 13 - прочие Gastropo-da, 14 - Orthocerida, 15 - Discosorida, 16 - Oncocerida, 17 - Ammonoidea, 18 - прочие Ce-phalopoda, 19 - прочие Mollusca, 20 - Odontopleurida, 21 - Proetida, 22 - Phacopida, 23 - прочие Trilobita, 24 - Palaeocopida, 25 - Metacopida, 26 - Podocopida, 27 - прочие Ostraco-da, 28 - прочие Arthropoda, 29 - Cystoporata, 30 - Trepostomata, 31 - Fenestrata, 32 – про-чие Bryozoa, 33 - Inarticulata, 34 - Orthida, 35 - Strophomenida, 36 - Pentamerida, 37 – Rhynchonellida, 38 - Spiriferida, 39 - Atrypida, 40 - Terebratulida, 41 - Crinoidea, 42 – про-чие "Pelmatozoa", 43 - "Eleutherozoa", 44 - Graptolithina, 45 - Conodonta, 46 – Cephalaspi-domorphi, 47 - Placodermi, 48 - Chondrichthyes, 49 - Acanthodii, 50 - Osteichthyes
<![if !vml]> <![endif]>
<![endif]>
Рис. 14. Приращения числа родов в основных крупных группах морских живот-ных в карбоне и перми; а - турне, б - визе, в - серпуховский век, г - башкирский век, д - московский век, е - поздний карбон, ж - ассельский век, з - сакмарский век, и - ленерд, к - гвадалупий, л - джульфинский и дорашамский века. Основные группы слева напра-во: 1 - Radiolaria, 2 - Foraminifera, 3 - Permosphincta, 4 - прочие Porifera, 5 - Tabulata, 6 - Rugosa, 7 - прочие Coelenterata, 8 - "Vermes", 9 - Problematica, 10 - Pterioida, 11 - прочие Bivalvia, 12 - Archaeogastropoda, 13 - прочие Gastropoda, 14 - Orthocerida, 15 - Nautilida, 16 - Goniatitida, 17 - прочие Cephalopoda, 18 - прочие Mollusca, 19 - Trilobita, 20 – Mala-costraca, 21 - Palaeocopida, 22 - Podocopida, 23 - прочие Ostracoda, 24 - прочие Arthropo-da, 25 - Cystoporata, 26 - Trepostomata, 27 - Fenestrata, 28 - прочие Bryozoa, 29 – Stropho-menida, 30 - Rhynchonellida, 31 - Spiriferida, 32 - прочие Brachiopoda, 33 - Monobathrida, 34 - Cladida, 35 - прочие Crinoidea, 36 - прочие "Pelmatozoa", 37 - "Eleutherozoa", 38 – Graptolithina, 39 - Conodonta, 40 - Pisces
<![if !vml]> <![endif]>
<![endif]>
Рис. 15. Приращения числа родов в основных крупных группах морских живот-ных в триасе; а - инд, б - оленек, в - анизий, г - ладин, д - карний, е - норий, ж - рэт. Основные группы слева направо: 1 - Spumellaria, 2 - Nassellaria, 3 - Foraminifera, 4 – Po-rifera, 5 - Scleractinia, 6 - прочие Coelenterata, 7 - Polychaeta, 8 - Pterioida, 9 - прочие Bi-valvia, 10 - Archaeogastropoda, 11 - Mesogastropoda, 12 - прочие Gastropoda, 13 – Nautili-da, 14 - Ceratitida, 15 - прочие Cephalopoda, 16 - Ostracoda, 17 - прочие Arthropoda, 18 - Bryozoa, 19 - Rhynchonellida, 20 - Spiriferida, 21 - Terebratulida, 22 - прочие Brachiopoda, 23 - Crinoidea, 24 - Holothuroidea, 25 - прочие "Eleutherozoa", 26 - Conodonta, 27 – Chondrichthyes, 28 – Reptilia
даннном случае регулирование может быть внешним по отношению к био-те. Для морских животных одним из лимитирующих факторов может быть общая площадь мелководных местообитаний, связанная с крупными тран-сгрессивно-регресивными циклами (Newell, 1967).
Высокая согласованность в изменениях разнообразия основных крупных групп выражается в отсутствии явного доминирования прираще-ний в каких-либо отдельных группах в общей структуре каждой из диа-грамм; вклад каждой группы в общее приращение разнообразия оказыва-ется более или менее соразмерным вкладу других групп. Однако, имеются исключения. Наиболее очевидным из них является раннекембрийский максимум разнообразия, полностью зависящий от изменений числа родов и семейств археоциат (рис. 1, 5, 11, г, д). В этом случае можно предпола-гать сильное влияние существенно комбинативной системы археоциат (Ро-занов, 1973) с большим количеством монотипных семейств и родов (Sep-koski, 1979; Дмитриев, 1989). Другой случай относится к гжельскому веку, когда общее снижение числа родов (рис. 1, 5, 14, е) определяется вымира-нием криноидей (по семействам вымирание в конце карбона распределяет-ся по крупным группам более равномерно).
По всей совокупности диаграмм не удается выявить какие-либо спе-циальные черты “массовых вымираний”, выделяющие их из общего хода
<![if !vml]> <![endif]>
<![endif]>
Рис. 16. Приращения числа родов в основных крупных группах морских животных в юре; а - геттанг, б - синемюр, в - плинсбах, г - тоар, д - аален, е - байос, ж - бат, з - келловей, и - оксфорд, к - кимеридж, л - титон. Основные группы слева направо: 1 - Spumellaria, 2 - Nassellaria, 3 - Foraminifera, 4 - Demospongia, 5 - Calcarea, 6 - Hexactinellida, 7 - Coelenterata, 8 - Annelida, 9 - Pterioida, 10 - Trigonioida, 11 - Veneroida, 12 - прочие Bivalvia, 13 - Archaeogastropoda, 14 - Mesogastropoda, 15 - прочие Gastropoda, 16 - Ammonoidea, 17 - прочие "Ectocochlia", 18 - Coleoidea, 19 - Podocopida, 20 - прочие Ostracoda, 21 - прочие Arthropoda, 22 - Bryozoa, 23 - Rhynchonellida, 24 - Terebratulida, 25 - прочие Brachiopoda, 26 - "Pelmatozoa", 27 - Echinoidea, 28 - прочие "Eleutherozoa", 29 - Chondrichthyes, 30 - Osteichthyes, 31 - Reptilia
последовательных перемещений диаграммы из одной полуплоскости в другую. Исключение составляет только маастрихтское вымирание, хотя в кампане уже проявляется тенденция к перемещению диаграммы вниз. По-степенность процесса, отображаемая диаграммами, может быть связана с влиянием эффекта Сигнора-Липпса (Signor, Lipps, 1982), но только в тех случаях, когда скорость вымирания наибольшая перед минимумом разно-образия, как указывалось выше (раздел по морским животным). Более рез-кими, чем вымирания, обычно оказываются переходы к новой диверсифи-кации после минимумов разнообразия.
Единообразие общей картины коллективного поведения всей сово-купности крупных таксонов не дает возможности легко выявить какие-ли-бо группы, ведущие себя особым образом вне общей тенденции. Это ука-зывает на отсутствие или слабое проявление различий групп по их реак-ции на общий рост или уменьшение разнообразия, то есть разделение фо-нового и “массовых” вымираний по их таксономической селективности на диаграммах не выражено.
Диаграммы приращений позволяют положительно ответить на воп-рос об обоснованости суммирования данных по морфологически удален-
<![if !vml]>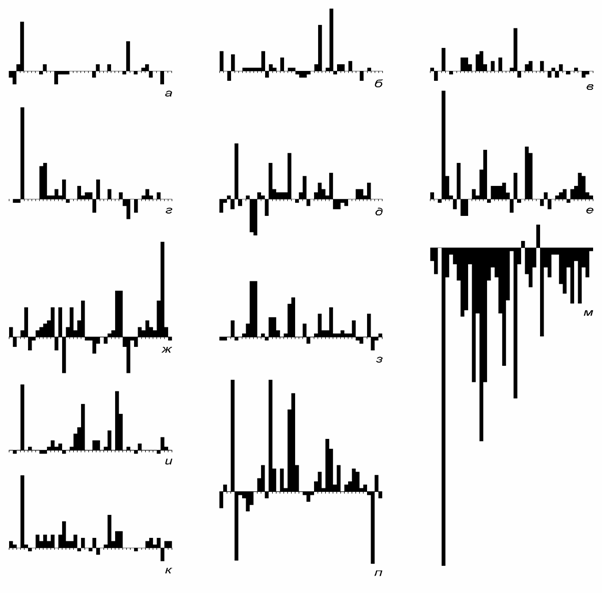 <![endif]>
<![endif]>
Рис. 17. Приращения числа родов в основных крупных группах морских живот-ных в мелу; а - берриас, б - валанжин, в - готерив, г - баррем, д - апт, е - альб, ж – сено-ман, з - турон, и - коньяк, к - сантон, л - кампан, м - маастрихт. Основные группы слева направо: 1 - Spumellaria, 2 - Nassellaria, 3 - Ciliata, 4 - Foraminifera, 5 - Lithistida, 6 – про-чие Demospongia, 7 - Calcarea, 8 - Hexactinellida, 9 - Scleractinia, 10 - прочие Coelenterata, 11 - Annelida, 12 - Pterioida, 13 - Trigonioida, 14 - Veneroida, 15 - Hippuritoida, 16 – Phola-domyoida, 17 - прочие Bivalvia, 18 - Cephalaspidea, 19 - Mesogastropoda, 20 – Neogastro-poda, 21 - прочие Gastropoda, 22 - Nautilida, 23 - Ammonoidea, 24 - Coleoidea, 25 - прочие Mollusca, 26 - Malacostraca, 27 - Ostracoda, 28 - прочие Arthropoda, 29 - Cheilostomata, 30 - прочие Bryozoa, 31 - Rhynchonellida, 32 - Terebratulida, 33 - прочие Brachiopoda, 34 - "Pelmatozoa", 35 - Cassiduloida, 36 - Holasteroida, 37 - Spatangoida, 38 - прочие Echinoi-dea, 39 - прочие "Eleutherozoa", 40 - Chondrichthyes, 41 - Osteichthyes, 42 - Reptilia, 43 - Aves
<![if !vml]>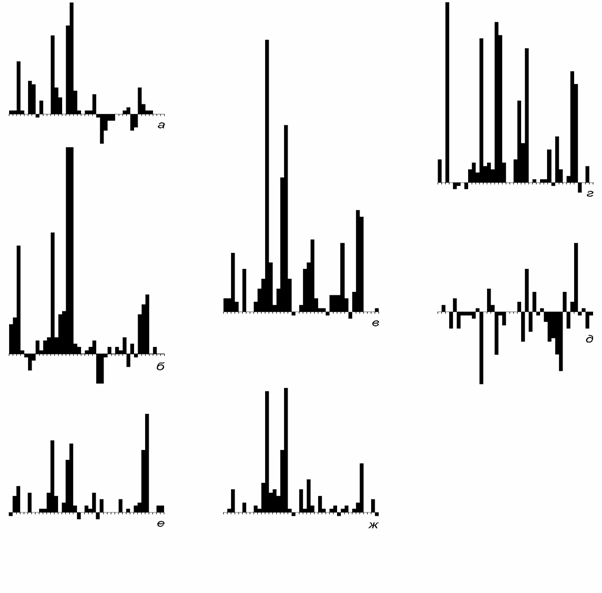 <![endif]>
<![endif]>
Рис. 18. Приращения числа родов в основных крупных группах морских живот-ных в палеогене; а - даний, б - палеоцен, в - ипр, г - лютет, д - поздний эоцен, е – рю-пель, ж - хат. Основные группы слева направо: 1 - Spumellaria, 2 - Nassellaria, 3 – Fora-minifera, 4 - прочие Protozoa, 5 - Porifera, 6 - Scleractinia, 7 - прочие Coelenterata, 8 – An-nelida, 9 - Nuculoida, 10 - Arcoida, 11 - Pterioida, 12 - Veneroida, 13 - прочие Bivalvia, 14 - Archaeogastropoda, 15 - Heterostrophia, 16 - Mesogastropoda, 17 - Neogastropoda, 18 – прочие Gastropoda, 19 - Cephalopoda, 20 - прочие Mollusca, 21 - Cirripedia, 22 – Malacos-traca, 23 - Ostracoda, 24 - Cheilostomata, 25 - прочие Bryozoa, 26 - Terebratulida, 27 – про-чие Brachiopoda, 28 - "Pelmatozoa", 29 - Temnopleuroida, 30 - Clypeasteroida, 31 – Cassidu-loida, 32 - Spatangoida, 33 - прочие Echinoidea, 34 - прочие "Eleutherozoa", 35 – Chond-richthyes, 36 - Perciformes, 37 - прочие Osteichthyes, 38 - Reptilia, 39 - Aves, 40 - Cetacea, 41 - прочие Mammalia
<![if !vml]> <![endif]>
<![endif]>
Рис. 19. Приращения числа родов в основных крупных группах морских живот-ных в неогене и антропогене; а-в - миоцен: а - ранний, б - средний, в - поздний, г – плиоцен, д - антропоген. Основные группы как на рис. 18
ным группам организмов на одном графике. Коллективность изменений разнообразия обеспечивает адекватность качественных характеристик суммарного графика - уже упоминавшихся выше интервалов монотонно-сти и экстремумов. График действительно отображает общие изменения разнообразия, а не разрозненные выбросы в отдельных группах. Однако, примеры археоциат и криноидей показывают, что отсутствие таких случа-ев не гарантировано, и контроль с помощью диаграмм всегда желателен, хотя не всегда возможен. Так, коллективное поведение не удалось убеди-тельно выявить для семейств неморских водных животных (Дмитриев и др., 1995, см. ниже главу о неморской водной биоте) из-за малого числа основных групп.
Диаграммы, как и приведенные выше графики, показывают поведе-ние реально изученного (актуального) разнообразия организмов, но в от-личие от графиков, они позволяют обосновать экстраполяцию за его пре-делы. Коллективность изменений в известной части разнообразия свиде-тельствует о сходных изменениях и в недоступной для изучения его части, то есть кривая числа таксонов качественно характеризует изменения всего существовавшего в прошлом разнообразия морских животных (но см. раз-дел “Полное разнообразие”). Отсюда же следует объяснение устойчивости общей формы кривых разнообразия в процессе изучения. Устойчивость кривых на разных стадиях исследования палеонтологического материала сама по себе также позволяет предсказать результаты последующего изу-чения, но в этом случае может быть дан только осторожный прогноз для той же совокупности групп ископаемых организмов.
С позиций первой интерпретации кривых разнообразия для числа таксонов они основаны, как обсуждено выше, на предположении о сред-нем морфологическом объеме таксона определенного ранга как общей единице разнообразия для всех суммированных групп, что противоречит обычным интуитивным представлениям систематиков. Вопрос принципиа-лен, так как график для одной крупной группы также суммирует данные о весьма различных составляющих группу таксонах. Средний объем таксо-нов может оказаться средним некоторого полимодального распределения с очень большой дисперсией, суммирование в таком случае приводит к не-явному введению неизвестных весовых коэффициентов. При постепенной смене основных доминирующих групп возможен тренд среднего объема таксонов, искажающий общую форму кривой числа таксонов по сравне-нию с действительной картиной изменений морфологического разнообра-зия в геологическом прошлом. Еще более неудобны резкие изменения ос-новного группового состава.
Применительно к диаграммам приращений эти соображения означа-ют, что при переходе к мере разнообразия величина приращений для раз-ных групп может непредсказуемым образом измениться, хотя вряд ли можно опасаться серьезных изменений картины в целом. Все же желатель-но, чтобы подобные предположения получали эмпирическое подтвержде-ние.
Интерпретация кривых разнообразия с помощью диаграмм прираще-ний чисто качественная. Она не дает обоснования применению количест-венных показателей диверсификации. В этом отношении первая интерпре-тация является более перспективной (как видно на примере по морским животным, иллюстрирующем, в частности, необходимость изучения ско-ростей), и в следующих разделах развивается количественный подход к анализу изменений разнообразия. Вторая интерпретация, кроме того, об-наруживает нежелательную зависимость обоснования кривых разнообра-зия от особенностей изучаемого процесса. Если коллективность измене-ний отсутствует или не продемонстрирована, смысл суммарной кривой становится неопределенным.
Обе интерпретации кривых разнообразия выявляют многочисленные проблемы, возникающие при использовании таксона как единицы морфо-логического разнообразия. Без таксонов обойтись не удается, но цена это-го посредничества кажется слишком высокой. В перспективе можно ожи-дать перехода к применению мер морфологического разнообразия.
Признаки. Наблюдаемый носитель разнообразия - не таксон, а эк-земпляр со свойственными ему изученными элементарными признаками. Определяющими для оценки разнообразия являются различия между эк-земплярами; их морфологическая сложность не имеет значения. В прост-ранстве признаков реально наблюденное (актуальное) морфологическое разнообразие отображается множеством точек, в котором точке соответст-вует экземпляр или группа экземпляров с общей характеристикой, как это обычно бывает в палеонтологических описаниях видов. Непрерывная из-менчивость признака в группе однородных по остальным признакам эк-земпляров разбивается на классовые интервалы, и группа отображается несколькими точками. Материал, представляющий исходные сведения в подходящей форме, частично накоплен в смежной области - в результате подготовки данных для обработки методами нумерической (фенетичес-кой) таксономии (для оценки разнообразия может потребоваться некото-рая детализация). Это создает предпосылки для применения мер морфоло-гического разнообразия. (В последние годы фенетическая таксономия в значительной мере уступила место кладистским методам).
Мера. Мерой морфологического разнообразия может служить сумма всех таксономических расстояний между точками (в математике - мера Ле-бега). Простейшее таксономическое расстояние - квадрат эвклидова рас-стояния. Когда в матрице признаков используются только 0 и 1, оно равно числу признаков, различающих соответствующие экземпляры или группы экземпляров. Для k экземпляров (групп экземпляров) простейшая мера морфологического разнообразия - сумма чисел различающихся признаков для k(k-1)/2 попарных сравнений экземпляров (групп). Если k - число всех экземпляров (групп), известных в составе таксона, то мера разнообразия равна морфологическому объему таксона. В литературе по фенетической таксономии (Sneath, Sokal, 1973; Quicke, 1996) обсуждаются различные ко-эффициенты сходства и связанные с ними таксономические расстояния. Все они могут быть применены для измерения морфологического разнооб-разия (многие коэффициенты эквивалентны друг другу - Песенко, 1982). Возможно использование мер, учитывающих морфологическую слож-ность, так как при большом числе признаков более вероятно обнаружение различий и появление новых точек с соответствующим увеличением объе-ма таксонов. В общем виде идея об использовании мер в пространстве признаков высказывалась и ранее, но не нашла применения из-за сложнос-тей практической реализации (Foote, 1993).
При использовании меры морфологического разнообразия величины в основном равенстве (1) приобретают следующий смысл: N+(t) - мера множества точек, соответствующих экземплярам (группам экземпляров), найденным в отложениях с возрастом древнее t; N-(t) - мера множества точек, соответствующих последним (наиболее молодым) находкам экземп-ляров, отличия которых от каких-либо более древних не удается выявить и находки которых древнее t; N(t) - мгновенное разнообразие в момент t, вы-раженное в единицах используемой меры.
Виды. Необходимость рассматривать во времени группы экземпля-ров, внутри которых они принимаются неотличимыми, означает использо-вание классификации видового и внутривидового уровня. Внутривидовые группировки, как правило, не будут выходить за пределы одного века, больший интерес они могут представлять в сравнительно редких случаях очень полно изученных региональных последовательностей. Во многих описаниях (возможно в большинстве) виды характеризуются в целом. Тог-да можно зафиксировать лишь первые и последние находки представите-лей вида, который в таком случае оказывается элементарной единицей анализа с мерой 0. Это не означает какой-либо фундаментальной роли ви-да (в контексте обсуждаемой проблематики) и зависит лишь от обилия коллекционного материала и глубины его изучения.
Единица измерения. Полного исключения таксонов в описываемом способе измерения не происходит, но их роль становится иной. Мера раз-нообразия может быть применена и к очень дробным внутривидовым еди-ницам, ограничение уровнем вида предлагается здесь только из сугубо практических соображений. Единицей измерения становится не таксон, а некоторое различие между теми или иными группами экземпляров, в част-ности, видами. Измеряется различие между комбинациями признаков, и основой работы становится признак. Признаки в процессе изучения более консервативны, чем таксоны. Привлечение нового признака в результате усовершенствования морфологических знаний - более редкое событие, чем изменение в системе. Число признаков многократно меньше числа таксонов, и в этом смысле они более элементарны, исследование разнооб-разия в целом переходит на следующий, теперь уже предельный уровень детальности. Признак в данном случае рассматривается как элементарная единица имеющихся знаний, а мера морфологического разнообразия как количественное описание знаний. Число таксонов также описывает накоп-ленные знания, но в этом случае единица не элементарна. Морфологичес-кая природа видов в палеонтологии определяет принципиальную обосно-ванность применения меры морфологического разнообразия.
Аддитивность. Другой аспект использования системы связан с ад-дитивностью меры. Объем таксона высокого ранга, вычисленный без уче-та его разделения на подчиненные таксоны, больше суммы объемов под-чиненных таксонов, так как в первом случае суммируются все таксономи-ческие расстояния, а во втором только внутри подчиненных таксонов. Полная мера морфологического разнообразия всей совокупности сильно удаленных таксонов будет включать сравнения, которые не учитываются в обычной работе систематиков. Чтобы получаемая картина более или менее удовлетворительно описывала результаты повседневной практики, следует отсечь неиспользуемые сравнения. Подходящей мерой во многих случаях, возможно, будет сумма объемов семейств (иногда надсемейств или отря-дов). При ограничении детальности анализа уровнем видов основой для работы становится матрица таксономических расстояний видов в пределах семейства, то есть обычный рабочий материал фенетической таксономии. Для полного исследования крупной группы требуется однотипное кодиро-вание признаков и использование единого показателя сходства для всех семейств. Отличие от работы систематика заключается, кроме отсутствия таксономической процедуры (в связи с другими целями исследования) и использования элементов уже имеющейся системы, в необходимости от-слеживать величины N+(t) и N-(t), тогда как при построении системы вре-менной аспект отсутствует.
Уточнение. Приведенное выше новое определение N+(t), N-(t) позво-ляет уточнить первую интерпретацию кривых разнообразия. В соответст-вии с ней графики N+(t), N-(t) для таксонов отображают накопленное число событий появления и вымирания, а N(t) - изменения морфологического разнообразия. Согласование одного с другим на уровне таксонов возмож-но только через константный средний объем таксона, что представляется слишком огрубленным, поскольку этот же средний объем приходится при-писывать таксонам в моменты их появления или вымирания. Переход к мере устраняет это осложнение.
Статистики. Применение меры морфологического разнообразия к изучению общей картины диверсификации ископаемых организмов воз-можно после предварительной подготовки обширного палеонтологическо-го материала. До того на основе меры и в связи с ней могут исследоваться некоторые частные вопросы. Так, перспективным направлением может быть статистика матриц признаков и сходства и объемов таксонов. В част-ности, представляет интерес распределение таксономических расстояний, которое часто соответствует распределению числа различающихся призна-ков (если таксономическое расстояние учитывает только число совпадаю-щих и различающихся признаков без отдельного учета совпадений отсут-ствия признака). В западной палеонтологической литературе в последние годы морфологическое разнообразие иногда сравнивается с помощью сум-мы дисперсий признаков (Foote, 1993). При одном и том же числе призна-ков (чаще всего в близкородственных группах или одной группе на разных временных срезах) и постоянном коэффициенте вариации матрицы приз-наков сумма пропорциональна среднему квадрату эвклидова расстояния, то есть (при кодировании признаков 0 и 1) среднему числу различающихся признаков. Для описания морфологического разнообразия, как оно опре-делено здесь, этого недостаточно. Кроме того, в данном случае могут силь-но влиять психологические характеристики таксономической процедуры.
Распределение объемов семейств определяется распределением так-сономических расстояний и распределением числа видов в семействе. Не-которые методы обработки данных в фенетической систематике включают вычисление среднего таксономического расстояния и, следовательно, объема, так что эти характеристики в ряде случаев могут быть уже извест-ны. Все три распределения отражают не только определенную структуру актуального разнообразия, но и результаты таксономического процесса, давая возможность исследования его с точки зрения особенностей работы систематика. Для двух крупных близких семейств с одним и тем же сред-ним таксономическим расстоянием квадратные корни морфологических объемов пропорциональны числу видов, то есть обиходному в системати-ке неформальному объему таксона.
Систематика. При большом k число возможных попарных сравне-ний растет примерно как k2/2. Вследствие этого создание системы или так-сономическая ревизия крупной группы могут быть чрезмерно трудоемки-ми. Естественным путем сокращения числа осуществляемых сравнений яв-ляется использование иерархической классификации (Беклемишев, 1994), которая возникает как способ работы при небезграничных возможностях классифицирующих субъектов (практический путь иерархизации через вы-деление типов позволяет еще более сократить число производимых срав-нений; в этом состоит таксономическая (не номенклатурная) роль типов).
Связь с уравнением (3). Эти замечания общего характера имеют от-ношение к уравнению (3) из раздела о разнообразии морских животных. Используя соотношение для числа сравнений Zi
Zi » aiNi2/2, (8)
где ai - доля осуществляемых сравнений, i - ранг таксона, от (3) (см. начало главы) можно перейти к равенству для числа сравнений Zi (и далее меры Mi). В данном случае принимается во внимание сокращение числа сравне-ний, связанное с иерархической классификацией. В пространстве призна-ков подмножество точек, соответствующих роду, семейству или отряду за-меняется точкой в центре подмножества. Мера уменьшенного множества точек
Mi = biZi,
где bi - среднее таксономическое расстояние, учитывающее признаки, ис-пользуемые для сравнения таксонов i-го ранга, и игнорирующее признаки, имеющие значение только внутри них. В качестве элементов разнообразия в этом случае рассматриваются таксоны i-го ранга, в отличие от описанно-го выше способа определения меры, подразумевающего наибольшую де-тальность анализа.
Подстановка в (3) из (8) дает равенство
Zi+1 = A1(B+lnZi)2r, (9)
где A1=ai+1A2/22r+1, B=ln2-lnai, то есть зависимость того же вида с дополни-тельным множителем при Zi. При стабильных bi, bi+1 величины Zi, Zi+1 мо-гут быть заменены на Mi, Mi+1.
В отсутствие теоретического обоснования зависимости (3) сближе-
ние ее с (9) является формально-спекулятивным. Тем не менее, удовлетво-рительная аппроксимация по (3) позволяет предположить примерное пос-тоянство величин ai, ai+1 для всего фанерозоя (кроме интервалов сущест-венного различия фактических и вычисленных величин), то есть однотип-ную структуру системы в аспекте доли возможных сравнений. Этот вывод не должен измениться при уточнении зависимости (3). Более точного соот-ветствия кривых, фактической и вычисленной по уравнению регрессии, можно искать, эмпирически изучая величины ai, определяющиеся главным образом статистическим распределением таксонов i+1-го ранга по числу составляющих их таксонов i-го ранга. В кембрии и тремадоке несоответст-вие кривых наиболее выражено, и для этого же времени актуальное разно-образие вполне очевидно отличается своеобразными изменениями вели-чин ai, ai+1 (рис. 10).
Дифференцирование (3) приводит к равенству
NilnNidNi+1 = rNi+1dNi,
выявляющему роль отношения Ni/Ni+1 - мгновенного среднего числа таксо-нов i-го ранга в составе таксонов i+1-го ранга, то есть важнейшей характе-ристики того же распределения.
Рассмотрение связи между мерами и числом таксонов разного ранга в геологической истории должно быть предметом отдельного исследова-ния. В частности, интересны общие черты изменения числа видов и меры морфологического разнообразия, которые следуют из известных измене-ний числа родов. Они могут быть предсказаны на основе уравнения (3), ес-ли оценить параметры по нескольким точкам в разных частях фанерозоя. Современные представления о фанерозойской диверсификации на уровне видов весьма противоречивы (Mosbrugger, 1992; Sepkoski, 1993a).
Pull of the Recent. На основе объема таксонов можно получить мате-риал к вопросу об описанном Д. Раупом (Raup, 1979) “притяжении совре-менности” (Pull of the Recent). Высокая по сравнению с ископаемым мате-риалом изученность современных фауны (и флоры) приводит к обнаруже-нию ископаемых представителей современных таксонов в тех случаях, когда без знания современных представителей они скорее всего были бы включены в состав каких-либо других таксонов. Наряду с несомненным ростом разнообразия многих крупных таксонов, в фаунистических спис-ках мезозоя и особенно кайнозоя могут присутствовать слабо обособлен-ные, небольшие, существующие ныне группы. В результате общее увели-чение числа таксонов в мезозое и кайнозое, неуклонно наблюдающееся в подавляющем большинстве крупных групп, не обязательно отражает соот-ветствующее увеличение морфологического разнообразия.
Свой вклад в мезокайнозойский рост вносят также реликтовые сов-ременные группы, их роль предлагал учитывать В. В. Жерихин (1978), ко-торый при анализе меловых семейств насекомых использовал представле-ние о “практически вымерших” семействах. Сходным образом влияют ред-кие находки ископаемых представителей в целом плохо сохраняющихся групп, представленных в современной биоте (Raup, 1979). Другой фактор - большая доступность более молодых отложений для изучения, имеет пре-имущественное отношение к проблеме полноты палеонтологических дан-ных, которая рассматривается ниже. Без применения меры выяснение дей-ствительного значения “притяжения современности” возможно, если спе-циально создать систему только ископаемых организмов, полностью игно-рируя современных, но трудно ожидать, что такая работа будет выполне-на. На основе меры морфологического разнообразия решение этого вопро-са, хотя бы частичное, может быть очень простым. Достаточно сравнить средний объем семейств и родов в некоторых крупных группах в отдален-ные друг от друга периоды, например, в триасе и неогене.
Темпы эволюции. Другое направление в использовании меры мор-фологического разнообразия, которое может представлять большой инте-рес, - изучение скоростей морфологической эволюции. Эта область огра-ничивается преимущественно исследованием скорости изменений отдель-ных или взаимосвязанных признаков. В последние годы все чаще приме-няется многомерный анализ (Fenster, Hecht, Sorhannus, 1991), но во всех случаях изучаются небольшие филогенетические последовательности. Ме-ра морфологического разнообразия дает способ количественно описывать изменения всего комплекса признаков в любых крупных группах организ-мов и их совокупностях. Кажется перспективным использование мер, учи-тывающих морфологическую сложность групп (Schopf et al., 1975).
Состояние морфологических знаний. Мера морфологического раз-нообразия и морфологический объем таксонов зависят от состояния мор-фологических знаний. При вводе в употребление новых, ранее не исполь-зовавшихся признаков увеличивается число столбцов матрицы признаков. Реже, когда некоторые признаки оказываются артефактами, оно может уменьшаться. При уточнении морфологии экземпляров меняются отдель-ные компоненты в строке. Число строк увеличивается при разделении группы экземпляров и уменьшается при объединении. Все изменения мат-рицы признаков могут влиять на меру. При постоянно продолжающемся изучении групп влияние этого фактора на кривые разнообразия невелико, если мера изменяется сходным образом в разных возрастных интервалах. Представляет интерес изучение меры в историческом аспекте, в ходе мор-фологических и таксономических исследований крупных групп. Это дало бы возможность оценить влияние морфологических знаний на изучение процессов диверсификации и одновременно исследовать некоторые осо-бенности таксономических решений.
Неоднородность методов выделения скелетных остатков и их изуче-ния (традиционное и химическое препарирование, световая и электронная оптика и т. д.), неполная сохранность остатков и их неполная изученность могут вносить значительную неопределенность, так как некоторым компо-нентам матрицы признаков не удается приписать определенное значение. Это может затруднить или даже сделать невозможным применение меры разнообразия к организмам, известным преимущественно по неполным ос-таткам, как наземные тетраподы, насекомые или растения. То же относит-ся к разным стадиям онтогенеза, например, водных насекомых. В подоб-ных случаях возможно применение меры для гомологичных структур и одинаковых стадий. Если в матрице признаков пробелов немного, неиз-вестным признакам может присваиваться значение от 0 до 1 в зависимости от частоты признака и корреляции его с другими признаками. В других случаях можно сократить число столбцов, огрубляя морфологическую ха-рактеристику до ее типичной для группы детальности. Применительно к изучению диверсификации организмов уточнение и детализация морфоло-гических знаний имеют своим следствием повышение точности оценок изменений разнообразия.
Погрешности и отношения
Кривая числа таксонов всегда точно описывает изменения этой вели-чины в одном из возможных вариантов системы, геохронологической шка-лы и корреляции. Это не гарантирует, что она, кроме того, отражает дейст-вительные изменения разнообразия. Прежде всего, кривая показывает из-менения реально изученного (или актуального) разнообразия, которое не совпадает с разнообразием, потенциально доступным для коллекциониро-вания и последующего изучения. В свою очередь, потенциальное разнооб-разие меньше полного захороненного разнообразия и тем более общего разнообразия, существовавшего в геологическом прошлом. Вопросы, от-носящиеся к этой области исследований разработаны недостаточно. Обыч-но ограничиваются непосредственным анализом графиков, как и было сде-лано выше в разделе о морских животных. В данном разделе погрешности и отношения рассматриваются на примере числа таксонов, но все выводы в той же степени относятся и к мере морфологического разнообразия.
Изученность. Ситуация в целом описывается отношениями
dt= X/X*,
где X обозначает одну из установленных по имеющимся данным величин DN+, DN- или Nt - установленное число таксонов из интервала продолжи-тельностью t; X* - та же величина при полной изученности существовав-шего разнообразия, нижний индекс означает, что данная величина харак-теризует временной интервал t. От dt следует отличать dt, которая описы-вает только отношение Nt/Nt* мгновенных разнообразий в момент t. При сопоставлении разных временных срезов t1, t2, ... изменения Nt верно ото-бражают изменения Nt*, если dti=dtj, то есть изученность одинакова (i, j - номера границ). Для сравнения временных интервалов t1, t2, ... условие dti=dtj недостаточно, о чем будет сказано ниже.
Доля вымерших таксонов. В условиях, когда неизвестна, возможно велика и неодинакова неизученная часть полного разнообразия и, кроме того, вследствие несовершенства временной шкалы затруднено использо-вание скоростей, кажется естественной популярность такого показателя, как доля (процент) вымерших таксонов. Повышение доли интерпретирует-ся как признак более интенсивного вымирания, вплоть до массовых выми-раний (Raup, Sepkoski, 1984; Sepkoski, Koch, 1996), которым ставятся в со-ответствие максимумы соответствующей кривой (Sepkoski, 1996). Пред-полагается, что доля вымерших (и появившихся) таксонов правильно хара-ктеризует именно тот временной интервал, для которого подсчитана, но это не так или не всегда так.
Доля вымерших таксонов R=DN-/(N0+DN+) увеличивается не только с возрастанием числа вымерших таксонов, но и с уменьшением числа общих таксонов со смежным более древним веком и числа появившихся таксо-нов. Поэтому при постепенно снижающемся разнообразии, как в среднем девоне и франском веке, доля вымерших таксонов может закономерно воз-растать даже при незначительном росте числа вымерших таксонов. Изме-нения родов морских животных в этом хронологическом интервале харак-теризуются следующими числами:
DN+ DN- N0 N0+DN+ R,%
Ейфель 607 688 1256 1863 37
Живет 479 704 1175 1654 43
Фран 488 735 950 1438 51
В живете доля вымерших родов возрастает на 7% по сравнению с эй-фелем, во франском веке еще на 8%, в обоих случаях на 19% по отноше-нию к своему предыдущему значению. В то же время число вымерших ро-дов растет соответственно только на 2 и 4%. Основной вклад в рост доли вымерших родов в живете и фране вносит существенное уменьшение зна-менателя N0 + DN+. Здесь ясно видна зависимость изменения доли от пре-дыдущей истории, то есть она не может служить специфической характе-ристикой данного хронологического интервала (в связи с наличием в зна-менателе слагаемого N0). Намного более удачна в этом отношении величи-на DN-/DN+, основная для обсуждавшихся выше графиков N(N+).
Зависимость доли от продолжительности веков. Более того, зави-симость изменений доли вымерших таксонов от окружения изучаемого ве-ка и других факторов - продолжительности веков и неравномерной изу-ченности фаун при возможных колебаниях скоростей появления и выми-рания, может приводить к противоречивым результатам. Так, максимум доли вымерших родов устанавливается в среднем миоцене (Sepkoski, 1996). По абсолютной величине (8,9%) он невелик, но на графике, приве-денном Сепкоски, ясно обособляется. Согласно подсчетам по его списку родов доля закономерно растет в 2 раза от позднего олигоцена и затем так же закономерно снижается в 5 раз к плейстоцену. Однако, на графиках числа родов (рис. 1,в, 5) и диаграммах приращений (рис. 19,б,в) нет сниже-ния разнообразия, но есть стабильный рост, замедляющийся по сравнению с ранним миоценом и не отличающийся от позднемиоценового. Скорость роста составляет 29.9 рода/млн. лет и в среднем и в позднем миоцене (здесь временная шкала не вызывает опасений), отношение DN-/DN+ равно 0.74 в среднем и 0.72 в позднем миоцене. Имеется явное противоречие: кривые не показывают среднемиоценового снижения разнообразия, но максимум доли вымерших родов обычно интерпретируется как индикатор существенного вымирания.
Можно проиллюстрировать один из возможных случаев возникнове-ния ложного максимума доли вымерших таксонов с помощью простого числового примера. Пусть на нижней границе некоторого временного ин-тервала имеется 100 таксонов и происходит линейный рост разнообразия в течение трех веков с продолжительностями 5, 13 и 4 при постоянных ско-ростях: появления - 12, вымирания -10. Процент вымерших таксонов сос-тавит последовательно 31, 49 и 22. Максимум доли получен в результате простого подбора подходящих продолжительностей. Согласно обычной трактовке в среднем веке - массовое вымирание.
Здесь нет прямой аналогии со среднемиоценовым максимумом, так как продолжительности трех частей миоцена соотносятся иначе (7.1, 5.4 и 5.9 млн. лет), а снижение доли вымерших родов от среднего миоцена к позднему (0.2%) незначимо. Пример показывает возможное влияние про-должительностей и необходимость учитывать его.
Корреляция вымираний с продолжительностью веков. Доля вы-мерших (и появившихся) таксонов имеет полностью определенный смысл, если только все вымершие в течение века таксоны вымерли одномоментно (и все появившиеся возникли также одномоментно), то есть не существует фонового вымирания (появления). Такая модель процесса свойственна раннему катастрофизму. Возврат к ней казался оправданным для вымира-ний после публикаций Д. Раупа и Дж. Сепкоски (Raup, Sepkoski, 1984; Raup, 1986), в которых сообщалось об отсутствии корреляции числа вы-мерших семейств и продолжительности веков. Характер кривых на рис. 3, 4 и их обсуждение в разделе о морских животных явно противоречат это-му результату. А. С. Алексеев (1984) рассмотрел материал по морским ро-дам и семействам с кампана по средний эоцен и пришел к выводу о досто-верной корреляции числа вымерших таксонов с продолжительностями ве-ков. То же подтверждается при подсчете по списку родов Сепкоски.
Коэффициенты корреляции подсчитывались по векам в отдельности для каждого из указывавшихся выше, при описании графиков для морских животных, продолжительных временных интервалов. За исключением вы-мирания в арениг-турнейском интервале, в котором продолжительность слишком многих веков определяется делением длительности крупных хро-нологических единиц на число составляющих их веков, везде получены невысокие, но значимые коэффициенты. Они следующие (в скобках – ко-эффициент корреляции для числа появившихся родов; знаком ! помечена значимость корреляции больше 0.99): арениг - турне 0.1 (0.5), визе – ле-нерд 0.70 (0.75), анизий - альб 0.57! (0.59!), даний - ныне 0.63 (0.77!), фане-розой в целом 0.35! (0.31!). Поскольку понятие о скорости вымирания (появления) и фоновой (средней) скорости не является бессмысленным, истолкование долей вымирания и появления (как и числа появившихся и вымерших таксонов) возможно при условии, что продолжительности ве-ков известны с достаточной точностью, но это условие не соблюдается.
Влияние неравномерной изученности. Другой случай возникнове-ния ложных максимумов доли вымерших таксонов может быть связан с неравномерной изученностью. Его также можно пояснить с помощью про-стого числового примера. Пример нарочито утрирован и характеризует предельный случай. Пусть сохранялось постоянное число таксонов равное 50 на протяжении трех одинаковых временных интервалов равных 10. Скорости появления и вымирания равны 1, число появившихся и вымер-ших таксонов - по 10, доли вымерших таксонов во всех трех интервалах - 16.7%. Для простоты предполагается отсутствие таксонов и появившихся, и вымерших в течение одного временного интервала. Допустим изучен-ность ранней и поздней фаун 100%, а среднему интервалу соответствует хиатус в хронологической шкале, то есть изученность равна 0. В этих ус-ловиях хронологическая шкала включает два века, причем вымирание всех вымерших в пропущенном интервале таксонов оказывается приписанным первому веку, а появление - второму. Тогда доля вымерших таксонов в первом веке - 33.3%, во втором - 14.3%, а на кривой разнообразия появит-ся ложный минимум на их границе. Если несколько смягчить условия при-мера и полагать изученность фауны среднего интервала 50%, доли теперь уже в трех веках составят последовательно 25, 10 и 16,6%, ложный мини-мум разнообразия также присутствует (в обоих случаях будет повышаться и наблюдаемая скорость вымирания в первом интервале).
Таким образом, доля вымерших (и появившихся) таксонов, исполь-зуемая как средство преодоления неопределенности, возникающей вслед-ствие неравномерной изученности фаун, оказывается сильно зависящей от этой же неравномерности, как и минимумы кривой разнообразия. Пример показывает также аналогию между сниженной палеонтологической изу-ченностью и хиатусом в хронологической шкале - таков же будет резуль-тат полного отсутствия палеонтологической характеристики в некотором стратиграфическом интервале. Различие состоит в том, что сниженная па-леонтологическая изученность есть погрешность собственно палеонтоло-гических данных, которая может быть уменьшена при получении и изуче-нии дополнительных коллекций, тогда как для устранения хиатуса в хро-нологической шкале необходим комплекс стратиграфических исследова-ний.
Влияние стратиграфической корреляции. Искажения, зависящие от стратиграфических ошибок, частично сходны с погрешностями палеон-тологической изученности. В результате неверных датировок отложений часть разнообразия переносится из одного века в другой. В нем dt может стать больше 1 для любого компонента X. Вследствие неравномерной па-леонтологической изученности то же может произойти с величиной DN- при “оттеснении” вымершего разнообразия из хуже изученного века в бо-лее древние и DN+ - появившегося разнообразия, - в более молодые, но dtt во всех случаях, DN+ в недостаточно изученном и предшест-вующих ему веках, DN- в недостаточно изученном и последующих веках; dt может быть различна у компонент X. Искажениям (в обоих случаях - при стратиграфических ошибках и неодинаковой изученности) подверже-ны также доли появившихся и вымерших таксонов и отношение DN-/DN+, кроме того возможно возникновение ложных минимумов разнообразия.
В современной общей шкале, как можно надеяться, не должно быть слишком грубых неточностей, но обобщение палеонтологических данных всегда основано на некоторой таблице стратиграфической корреляции, где идеальным изохронным границам веков ставятся в соответствие опреде-ленные уровни в регионах и конкретных разрезах. Обоснованность поло-жения границ в разных случаях весьма отличается, часто оно очень нео-пределенно. Множественность корреляционных таблиц и непрерывные стратиграфические дискуссии не оставляют сомнений в неизбежности не-верных датировок. Особенно часты они должны быть в пограничных час-тях стратонов. Возможна взаимная компенсация возникающих при этом искажений, но она не обязательна и вряд ли будет полной.
Хотя на изучение эволюции разнообразия влияет стратиграфическая изученность, этот аспект находится за пределами собственно палеонтоло-гии в отличие от палеонтологической изученности. Поскольку искажения, связанные со стратиграфическими неточностями всегда выражаются в не-верных датировках первых и последних находок, они могут быть миними-зированы при аккуратном датировании в списках исходного материала. При компиляции данных о распространении таксонов желательно избегать прямого заимствования датировок из литературы без выяснения их поло-жения в принятой для компиляции шкале. Не следует стремиться к точной хронологической локализации находок в сомнительных случаях, лучше указывать наименьший несомненный интервал, к которому находка отно-сится. Тогда при обработке исходных данных такие датировки могут быть распределены по более дробным хронологическим интервалам, как указа-но выше (см. раздел “Первичная обработка данных”). Оценка величины искажений, зависящих от состояния стратиграфии, может быть осущест-влена, повидимому, только при сравнительном изучении картин эволюции разнообразия, получаемых на основе альтернативных стратиграфических корреляций.
Альтернатива долям. В отличие от доли вымерших и появившихся таксонов, отношение DN-/DN+ теоретически, для идеального случая высо-кой равномерной изученности может рассматриваться как выборочная ста-тистическая характеристика века и будет достоверным и достаточно точ-ным при обычных для веков величинах в несколько сот вымираний и поя-влений родов морских животных (семейств в большинстве случаев нес-колько десятков). При росте числа родов в ходе дальнейшего изучения су-щественных изменений DN-/DN+ в этом случае не должно происходить. По своему содержанию отношение идентично предложенному ранее В.В. Же-рихиным (1978) индексу когерентности (DN+- DN-)/(DN++DN-), но (как и этот индекс - Расницын, 1988) не взвешено по масштабу событий, что за-трудняет его изолированное применение. На графике N(N+) оно определя-ет угол наклона и хорошо вписывается в общую картину изменений разно-образия. В частности, должны быть очень наглядны эпизоды предполагае-мых катастрофических вымираний, так как при чистом вымирании (без появления) соответствующий отрезок графика становится вертикальным.
Статистические свойства отношения DN-/DN+ (и других) отличаются некоторым своеобразием. Совокупность таксонов определенного ранга, известных из отложений того или иного яруса, может составлять весьма значительную часть всего способного захорониться разнообразия древних скелетных организмов (при оценке его числом таксонов этого же ранга), в отличие от обычных выборок статистики, малых по сравнению с генераль-ной совокупностью. Поэтому высокая представительность материала во многих случаях обеспечена. С другой стороны, исходный для подсчетов список уравнивает таксоны, тогда как вероятность их присутствия в спи-ске изначально неодинакова. Кроме того, с позиций меры морфологичес-кого разнообразия, список игнорирует дисперсию объема таксонов.
Отношение DN-/DN+ более однородно по характеру исходного мате-риала, чем доли вымерших или появившихся таксонов, так как использует только фактические находки первых и последних представителей таксо-нов. В случае доли вымерших (появившихся) таксонов знаменатель вклю-чает также все таксоны, существование которых в течение века устанавли-вается не непосредственно, а по находкам только в более древних и более молодых отложениях (то же относится к удельным скоростям появления и вымирания). В этом отношении число таксонов в веке отличается и от мгновенного разнообразия, отнесенного к границе хронологического ин-тервала. Последнее учитывает только находки представителей таксонов ниже и выше границы.
В реальном иссследовании погрешности отношения DN-/DN+ могут быть значительными в связи с неравномерной палеонтологической изучен-ностью и ошибками стратиграфической корреляции, вплоть до полной не-возможности его применения.
Пригодность данных. Требование dti=dtj для мгновенных разнообра-зий при переходе к временным интервалам, характеризуемым изученнос-тями dt, замещается другим условием. Если в нескольких веках существо-вало без изменений крупное надсемейство с двумя равными семействами, причем в отложениях выше границы с возрастом t встречены все предста-вители одного из них и только они, а ниже нее столь же полно представле-но другое скемейство, то dt = 0.5 для обеих фаун, эмпирическая доля вы-мерших к моменту t 100%, как и появившихся после него, dt составит для момента t и ближайших к нему границ 0.5, 0, 0.5, минимум разнообразия также неизбежен. Поэтому должно быть dti=dtj не только для всей изучае-мой фауны, но и в отдельности для каждого из составляющих ее таксонов при одинаковой изученности каждого из них. Если для всей совокупности групп dt<<1, это усиленное условие заведомо неосуществимо, а при dt=1 выполняется всегда. Точное условие пригодности данных dt=dt=d=1, но полная и равномерная по всей хронологической шкале изученность всех групп - слишком суровое требование к палеонтологическому материалу. На практике достаточно, чтобы общая изученность была близка к 1 и не слишком отличалась для смежных границ, то есть менялась медленно (аналогично такому же условию для среднего объема таксонов в первой интерпретации кривых разнообразия). Например, если фауна начала века известна с dt = 0.85, а в конце его dt = 0.88, то отношение разнообразий на границах века определяется с относительной ошибкой 3.5%. Однако, отно-сительная ошибка доли вымерших таксонов может быть неприемлемо ве-лика даже при высокой изученности, если все существовавшие, но не об-наруженные в первом из двух веков группы представлены во втором и на-оборот (предельный случай).
Репрезентативность. Предельный случай маловероятен, а абсолют-ная ошибка доли может быть не очень большой - здесь существен сам под-ход к этим вопросам. Часто используется оценка погрешности доли вы-мерших таксонов с помощью стандартного метода, основанного на бино-миальном распределении (Sepkoski, Koch, 1996). Считается, что доля вы-мерших (или появившихся) таксонов в их списке дает статистически обос-нованную оценку этой же доли в изучаемом древнем разнообразии. Но при неравномерной изученности обоих типов - по вертикали и между группа-ми, возможна и может быть обычной ситуация, когда dt для общего числа таксонов и для числа вымерших (появившихся) таксонов различны. Тогда доля вымерших (появившихся) таксонов характеризует не природное раз-нообразие, а только сам список таксонов, который в этом случае не имеет статуса подходящей для исследования выборки. Ошибка доли (как и лю-бых других отношений и коэффициентов) теряет свой смысл, так как изу-чаемой генеральной совокупностью фактически оказывается список таксо-нов, а доля - генеральной долей в этой совокупности. Происходит неявная подмена объекта изучения (ниже будет рассмотрен другой похожий слу-чай). Более детальный анализ погрешностей возможен при учете вероят-ностей присутствия таксонов в фаунистическом списке.
Фактическое и интерполированное присутствие. Некоторая кор-ректировка коэффициентов и отношений, включающих общее число так-сонов, возможна при учете только таксонов, фактически присутствующих в отложениях того или иного века, и исключении таксонов, существование которых устанавливается только по находкам в более древних и более мо-лодых отложениях. Такой подход устраняет неоднородность используемо-го фаунистического списка со смешением реально найденных и не пред-ставленных в каменном материале таксонов и зависящие от нее артефакты, как в случае с долей вымерших таксонов. Точно также оценка вероятнос-тей появления и вымирания (основной характеристики процесса) будет бо-лее адекватной, если для нее используются только фактически найденные таксоны. Однако, во всех сводках с датировками только первых и послед-них находок необходимая информация отсутствует. Кроме того, такое уточнение количественных оценок не устраняет погрешностей, зависящих от неоднородной изученности.
Примат кривой разнообразия. Если d»1 список таксонов исчерпы-вает древнее разнообразие. Тогда основным средством изучения диверси-фикации будет сама кривая разнообразия как таковая, любые отношения становятся излишними, хотя могут использоваться для дополнительного более наглядного иллюстрирования особенностей процесса. При неполной изученности никакие отношения не свободны от различных искажений, а их интерпретация оказывается неоднозначной.
Кривая разнообразия в целом менее чувствительна к неполной изу-ченности. чем число или доля появившихся и вымерших таксонов. Так, при изученностях dt на смежных границах 0.30 и 0.35 относительная пог-решность будет 14%, но при этом отсутствует способ определения dt и оценка достоверности результатов невозможна. Это может быть сделано только при изученностях, приближающихся к 1 (см. ниже). Все изложен-ное является конкретизацией применительно к палеонтологическому мате-риалу простого факта: если имеются основания сомневаться в статистичес-кой репрезентативности выборки (неоднородная изученность исключает репрезентативность), никакие выборочные показатели неприменимы и ис-следование может быть основано только на изучении самой генеральной совокупности. Стремление палеонтологов составить как можно более пол-ный список таксонов и делать выводы, обобщая данные по всему списку, свидетельствует именно об оправданном недоверии к выборочным харак-теристикам.
Часто предполагают, что изученность много меньше 1 для всех групп вместе, непостоянна и различна для разных групп. Тогда первую ин-терпретацию кривых разнообразия следует признать нереалистичной и ограничиться второй. В этом случае придется отказаться от числовых оце-нок и использовать только осторожный качественный анализ направлен-ности процесса, имея ввиду возможность артефактов, порождаемых нерав-номерной и неполной изученностью.
Повышение ранга таксонов. Использование количественных хара-ктеристик подразумевает достаточно высокую изученность. Возможный способ увеличить ее - повышение ранга таксонов. Имеются сомнения в пригодности данных по видам, тогда как роды и семейства признаются подходящими (Шиманский, Соловьев, 1982). Смысл этого мнения очеви-ден: когда зафиксировано присутствие семейства в некотором возрастном интервале (даже если его представители найдены только в более древних и более молодых отложениях), любое число последующих находок предста-вителей семейства в этом интервале не меняет изученности на уровне се-мейств. Компьютерное моделирование (Lasker, 1978) показывает, как не-полнота изученности на низших таксономических уровнях может быть скомпенсирована при повышении ранга таксонов. Работа с высшими так-сонами связана с потерей детальности описания процесса, обсуждавшейся в первой интерпретации кривых разнообразия (регистрация присутствия семейства в нескольких последовательных временных интервалах ничего не говорит об изменениях в них числа родов или видов данного семейст-ва). Сама первая интерпретация при неполной изученности оказывается, кроме того, неприложимой к конкретному материалу: попытка применить меру разнообразия требует во всяком случае не меньшей изученности, чем работа на уровне видов. (Отсутствие детальных глобальных обобщений по многим группам вместе для видов в течение всего фанерозоя на практике определяется не общими соображениями пригодности данных, а отсутст-вием пока-что подготовленного должным образом материала).
Исключение непригодного материала. Еще одна возможность улучшить исходные данные - отбор достаточно полно изученных групп и отказ от использования известных лишь фрагментарно. Так поступил Сеп-коски (Sepkoski, 1996), приведя кривую доли вымерших родов с исключе-нием “плохо сохранившихся” групп. Этим способом удается несколько от-корректировать получаемые оценки, улучшая статистическую репрезента-тивность данных, но общая изученность уменьшается и полностью устра-нить погрешности невозможно.
Предварительное суждение о пригодности материала по той или иной группе можно получить, рассматривая распределение продолжитель-ностей существования родов (или семейств) в ее составе. Обычное распре-деление - показательное, обусловленное монофилетичностью родов (се-мейств) в достаточно изученных группах. А. В. Марков и Е. Б. Наймарк (1994) показали, что объединяемые в “искусственные семейства” случай-ные группы родов характеризуются (как и должно быть из вероятностных соображений при разрыве связи по происхождению) равномерным распре-делением продолжительностей существования. Там же было продемонст-рировано равномерное распределение продолжительностей существования семейств правильных морских ежей. Как правило, такие случаи будут со-пряжены с фрагментарной изученностью, препятствующей формированию хорошо обоснованной системы и, равным образом, изучению диверсифи-кации группы. Другой способ отбраковки недостаточно изученных групп упоминается в следующем разделе (см. также Foote, Sepkoski, 1999).
Хиатусы. Некоторая компенсация неполноты исходных данных воз-можна при введении поправок на хиатусы в распространении таксонов. Случаи, когда полное распространение семейства не покрывается установ-ленным распространением составляющих его родов, сравнительно неред-ки. Хиатусы более часты в окрестностях минимумов разнообразия и харак-терны преимущественно для фрагментарно изученных групп. Это может приводить к парадоксальному установлению в моменты минимального разнообразия большего числа семейств, чем родов. Для отрядов и входя-щих в них семейств хиатусы выявляются реже, для семейств и родов, ро-дов и видов - чаще. В таких случаях можно использовать поправку, про-должая до середины хиатуса распространение наиболее молодого рода из встреченных до хиатуса и наиболее древнего из встреченных после хиату-са. Если принимается какая-либо филогенетическая гипотеза, связываю-щая разделенные хиатусом роды, поправка может быть более сложной и, возможно, более точной. Ниже будет указан более эффективный метод, уменьшающий влияние не вполне верных филогенетических реконструк-ций.
Значение интервальных изученностей. Адекватность кривой разнообразия (числа таксонов) прямо зависит от dt и косвенно от dt, так как примерное постоянство dt обеспечивается только при высоких изучен-ностях dt. В отличие от кривой разнообразия, графики N+(t), N-(t) непо-средственно зависят от соответствующих dt.
Классификация погрешностей. Обобщая изложенное выше, можно дать следующую систематизацию источников погрешностей:
1) ошибки стратиграфической корреляции;
2) точность временной (радиоизотопной) шкалы;
3) измерение разнообразия числом таксонов и неточности системы групп;
4) неравномерность палеонтологической изученности фаун разного возраста;
5) неоднородность изученности разных групп.
Комментарии к классификации. Первые две категории погрешнос-тей обусловлены факторами, не относящимися к палеонтологии в узком смысле. Некоторые способы уменьшить влияние погрешностей первой группы были указаны выше. Погрешности временной шкалы являются очень существенными при изучении скоростей и вероятностей появления и вымирания. Сравнения скоростей и вероятностей в одном веке от них не зависят. Графики N(N+) используют накопленное число появившихся так-сонов N+ в качестве собственного (внутреннего) времени процесса дивер-сификации. Отсюда следует принципиальная возможность широкого срав-нительного изучения скоростей диверсификации и вымирания с использо-ванием N+ для многих групп вместе в качестве независимой переменной. Перспективы и ограничения этого подхода не вполне ясны.
В разделе о первой интерпретации кривых разнообразия обсуждены недостатки числа таксонов как средства измерения разнообразия. В случае перехода к непосредственному использованию меры морфологического разнообразия погрешности, зависящие от использования числа таксонов и через него, опосредованно - с морфологической изученностью, заместятся погрешностями, прямо связанными с изученностью морфологии групп ор-ганизмов.
Имеется еще один тип погрешностей, зависящий от системы в ее связи с филогенией. Момент филетического вымирания отражается в спи-ске таксонов как вымирание предкового таксона и одновременное появле-ние нового таксона. Число таксонов на границах не изменится, если филе-тическое вымирание произошло в течение века, в противном случае оцен-ка разнообразия на границе, с которой совпало это событие, будет заниже-на. Число учитываемых появлений и вымираний увеличится в любом слу-чае, хотя реального вымирания и появления не было. Значение этого фак-тора зависит от относительной роли филетического вымирания и дивер-генции и может быть различным в разных крупных группах. Влияние фи-летического вымирания в целом в данной работе предполагается несу-щественным, так как в подавляющем большинстве известных автору слу-чаев устанавливается одновременное существование предкового и произ-водного от него таксона. Даже в тех нечастых случаях, когда удается де-тально проследить на популяционном уровне переход одного вида в дру-гой, крайние члены переходной популяции таксономически иногда отно-сят к разным видам (Левен, 1970), что определяется морфологической природой вида в палеонтологии.
Другой тип искажений имеет преимущественное отношение к исто-рии палеонтологии и стратиграфии и связан со специализацией исследо-вателей на изучении отложений определенного возраста, что в прошлом, сочетаясь с катастрофистскими представлениями о неизбежных крупных изменениях таксономического состава на рубежах единой шкалы, нередко приводило к появлению синонимов (горизонтальная классификация). В последние десятилетия в связи с более узкой таксономической специализа-цией палеонтологов и особым интересом к границам систем, определив-шим внимательное изучение пограничных отложений, синонимия по край-ней мере надвидовых названий, как можно надеяться, во многом выявлена и устранена.
Погрешности четвертого и пятого типа совместно устраняются при полной изученности. Для их выявления и оценки необходимо выяснение полноты палеонтологических данных. Погрешности пятого типа возника-ют при неравномерной изученности по вертикали некоторых групп из чис-ла составляющих всю изучаемую совокупность. Они сильно влияют на адекватность системы: при фрагментарной изученности в системе группы могут превалировать случайные объединения неродственных форм.
Применительно к количественному изучению полнота палеонтоло-гических данных может обсуждаться в двух аспектах:
1) полнота реально изученного или актуального разнообразия по от-ношению к потенциально доступному для изучения материалу;
2) соотношение с полным захороненным разнообразием.
Оба вопроса касаются уже не отображения имеющихся знаний (акту-альное разнообразие), а их отношения к той реальности, которую эти зна-ния должны описывать. Ниже обсуждаются два подхода к проблеме соот-ношения актуального и потенциального разнообразий, первый из них наз-ван здесь асимптотическим, второй - экстраполяционным. Асимптотичес-кий подход ориентирован на выяснение пригодности имеющихся данных, экстраполяционный преследует более далеко идущие цели улучшения ко-личественных оценок.
Принцип асимптотического метода иллюстрируют несколько пара-доксальные графики на рис. 20. По данным таблицы распространения ро-дов гастропод из работы О. В. Амитрова и др. (1981) построены графики двух типов для кампана (рис. 20, а, б), маастрихта (рис. 20, в, г), дания (рис. 20, д, е) и монса (рис. 24, ж, з). По горизонтальной оси всех графиков обозначены годы, по вертикальной - число родов N1 на нижней границе ве-ка (левые графики) и отношение числа родов N2/N1 на верхней границе ве-ка к числу родов на его нижней границе (правые графики). Ординаты то-чек на всех графиках соответствуют числам, получаемым при учете только родов, выделенных до года, обозначенного у горизонтальной оси. Даты первоописаний взяты из справочников “Treatise on invertebrate paleontolo-gy”, “Основы палеонтологии” и сводок В. Венца (Wenz, 1938-1944, Wenz, Zilch, 1960). Учитывались только морские гастроподы, материал в исход-ной таблице – глобальный. Все графики могут показаться лишенными ка-кого-либо смысла. Это не совсем так, но необходимы довольно подробные пояснения.
В отложениях, доступных для сбора палеонтологических коллек-ций, захоронено некоторое разнообразие остатков организмов, которое обозначено здесь как потенциальное разнообразие. Если потенциальное разнообразие достаточно полно представлено в имеющихся коллекциях, новые сборы не меняют сколько-нибудь существенно уже известного (или актуального) разнообразия, и увеличение числа родов связано преиму-щественно с таксономическими ревизиями. Происходит в основном уменьшение среднего объема родов, а на графиках отношений (рис. 20, справа) должна выявиться горизонтальная асимптота.
Чтобы избежать кажущегося противоречия: актуальное разнообразие есть число известных таксонов, а увеличение числа таксонов не приводит к изменению актуального разнообразия, под актуальным разнообразием как и выше (см. раздел о мере) понимается множество точек в пространст-ве признаков. Тогда потенциальное разнообразие - множество точек, соот-ветствующее полной изученности; актуальное разнообразие является его подмножеством. “Чистые” таксономические ревизии (без нового каменно-го материала и уточнения морфологии) приводят только к изменениям в разбиении актуального разнообразия по таксонам, не меняя его самого. Ес- мера морфологического разнообразия - сумма объемов семейств, то при
<![if !vml]> <![endif]>
<![endif]>
Рис. 20. Число родов (слева) морских гастропод на нижней границе веков конца мела и начала палеогена и отношение (справа) числа родов на верхней границе к числу родов на нижней границе в зависимости от времени первоописания родов: а, б – кам-пан, в, г - маастрихт, д, е - даний, ж, з - монс. Данные по Амитров и др., 1981. Поясне-ния в тексте
чистых ревизиях родов она останется той же.
Упоминавшееся выше повышение изученности при переходе на бо-лее высокие таксономические уровни (интуитивно ясное) определяется ве-роятностью попадания точек из дополнения актуального разнообразия до потенциального в те части пространства признаков, которые при сущест-вующем разбиении актуального разнообразия по таксонам находятся вну-три границ одного из таксонов. Вероятность выше при близости актуаль-ного и потенциального разнообразий и более крупном членении актуаль-ного разнообразия на таксоны. Последнее дает еще одно объяснение высо-кой устойчивости кривых разнообразия в процессе изучения древних фа-ун. Общей таксономической тенденцией является дробление таксонов, и на предшествующих этапах изучения большее несоответствие актуально-го и потенциального разнообразий компенсировалось делением актуально-го разнообразия на более крупные части.
Потенциальное разнообразие в процессе изучения не константно и зависит от изученности морфологии организмов (см. раздел о мере). Оно определяется списком признаков, учитываемых матрицей признаков. В этом и последующих разделах потенциальное разнообразие считается за-фиксированным на современном уровне морфологических знаний.
На рис. 20 состав и распространение родов приняты в соответствии с представлениями авторов исходной таблицы ко времени ее составления. Корректное изучение подразумевает учет изменений таких представлений в ходе познания группы, для чего необходимы специальные исторические исследования: сведения о распространении таксонов в разные моменты ис-тории изучения обобщаются на основе современной стратиграфической шкалы и корреляции (стратиграфический аспект истории изучения в дан-ном случае излишен). Такие исследования трудоемки и более доступны при наличии развитых компьютерных баз данных. Без учета истории изу-чения точки на графике занимают положение, к которому они перемеща-ются от некоторого исходного “правильного” положения по мере того, как понимание состава и распространения родов приближается к современно-му. Величина перемещения точек уменьшается слева направо: при прибли-жении к современному состоянию изученности область, в пределах кото-рой точки располагались изначально, постоянно сужается. (Самая левая часть всех графиков на рис. 20 является фиктивной - она соответствует ро-дам, первоначально выделенным в составе современной фауны и лишь значительно позже обнаруженным в ископаемом состоянии в кампане – монсе. При полном исследовании эта часть графиков исчезнет).
Протяженная горизонтальная асимптота на графиках отношений N2/N1 (рис. 20, б, з) дает основания надеяться, что она сохранится и при полном исследовании, а представления об изменениях разнообразия в те-чение века являются достаточно обоснованными. Это делает оправданным дальнейший переход к полному анализу. Отсутствие выраженной горизон-тальной асимптоты (рис. 20,г,е) позволяет прогнозировать дальнейшее из-менение N2/N1 в ходе последующих исследований. Полный анализ скорее всего окажется преждевременным. Увеличение отношения N2/N1 для дания и уменьшение для маастрихта свидетельствуют о еще не стабилизировав-шейся оценке глубины маастрихт-датского минимума разнообразия морс-ких гастропод.
Имеется механизм, обусловливающий тенденцию к появлению лож-ной горизонтальной асимптоты на графиках отношений N2/N1 при непол-ном исследовании. Последовательность отношений (точек) на таких гра-фиках формируется в результате постепенного увеличения учтенного чис-ла родов, которое продолжается до исчерпания всего используемого спис-ка родов. Отношения можно рассматривать как выборочную статистичес-кую характеристику, уточняющуюся при увеличении объема выборки. Ге-неральной совокупностью в этом случае будет весь массив имеющихся данных (список), ордината крайней правой точки соответствует отноше-нию N2/N1 в генеральной совокупности. Горизонтальная асимптота выя-вится, когда учтенное число родов станет репрезентативным для списка в целом; ордината асимптоты может быть никак не связана с отношением потенциальных разнообразий.
Возможное появление ложной асимптоты при неполном исследова-нии предполагает, что в этом случае действительная история исследования может стираться вследствие подмены объекта изучения. Вместо постепен-но увеличивающихся выборок из неизвестного потенциального разнообра-зия (научно-исторической последовательности актуальных разнообразий) составляются выборки из исходно фиксированного массива данных (сов-ременного актуального разнообразия). Но если при неполном исследова-нии история всегда затушевывается, все графики будут выходить на гори-зонтальную асимптоту. На рис. 20 такая асимптота проявилась для кампа-на и монса, но не маастрихта и дания, то есть основные черты истории изучения все же сохраняются. Это должно быть следствием того, что по крайней мере в правой части “правильного” графика положение точек не сильно отличается от их положения на рис. 20 и перемещение точек в ходе изучения ограничено здесь узкой зоной вблизи кривой. В этом случае на-дежность выводов даже при отсутствии полного исследования может быть более высокой. Без графиков для маастрихта и дания сохранение истории не могло бы выявиться.
Горизонтальная правая часть графика отношений N2/N1 не обязатель-но связана с высокой однородной изученностью, которая может быть на сравниваемых границах и разной, но длительно сохраняющейся, и сходно меняющейся. Наиболее ясный случай появления горизонтальной асимпто-ты - постепенное повышение и одновременно стихийное выравнивание изученностей dt сравниваемых мгновенных разнообразий. Для такого про-цесса можно получить и строгое формальное обоснование, но примени-тельно к эмпирическому материалу асимптотический метод в своей основе индуктивен, и результаты желательно контролировать другими способами.
Левые графики на рис. 20 показывают изменения в ходе исследова-ний числа родов на нижней границе веков, что позволяет более полно по-нимать результаты, показанные на правых графиках. Они также отличают-ся от “правильных” графиков, но здесь прямолинейная трактовка кажется менее опасной. Обращает на себя внимание горизонтальный участок спра-ва на кривой для монса (рис. 20, ж). Увеличения числа родов не происхо-дит, начиная примерно с 1940 года. Такая же картина характерна для тане-та и веков эоцена. Причины этого явления могут быть различными, от ста-билизации системы палеогеновых морских гастропод на уровне родов к 1940 году, вследствие практически полной изученности родов, до непол-ного учета родов, выделенных после 1940 года, вследствие отсутствия ми-ровых справочников родового уровня для многих групп гастропод после сводки Венца (Wenz, 1938-1944). Поскольку число родов с 1940 года ста-билизируется, отношения N2/N1 (рис. 20, з) на этом участке графика также не меняются, но кривая отношений горизонтальна не только для 40 лет по-сле 1940 года, но и еще 80 лет до того, в течение которых происходил пос-тоянный рост числа описанных родов.
Монский случай иллюстрирует необходимость разностороннего ана-лиза, в частности, использованная здесь ось времени изучения может ока-заться не самой удачной. Абсциссой может быть любая величина, моно-тонно растущая при накоплении коллекций, прежде всего это число экзем-пляров (особенно если изучается отношение мер разнообразия), затем раз-личные паллиативы: число видов или родов (если изучается отношение для таксонов более высокого ранга), число местонахождений, точек сбора, коллекций (К. Кох и Дж. Морган (Koch, Morgan, 1988) успешно использо-вали число коллекций для других целей - см. ниже) и, наконец, время изу-чения. В случаях длительно сохраняющегося отношения N2/N1 при росте числа родов можно дополнительно анализировать способ роста: если акту-альное разнообразие на уровне родов близко к потенциальному, новые ро-да выделяются на основе уже известных видов, или такие виды указыва-ются в их составе, или включаются в состав рода впоследствии. На уровне видов то же будет с ранее описанными в составе других видов экземпляра-ми.
При повышении изученности групп происходит постепенное вытес-нение новых таксонов, описываемых по материалу вновь поступающих коллекций, новыми таксонами, появляющимися в ходе таксономических ревизий и не включающих существенно нового коллекционного материа-ла. Этот признак изученности может использоваться для предварительной оценки пригодности данных (Дмитриев, 1978), а также оценки уже постро-енных графиков. Так, возрастание числа родов на границах кампана (рис. 20, а, в) при асимптотическом поведении отношения N2/N1 (рис. 20, б) мо-жет означать увеличение числа родов преимущественно в результате пере-смотра системы без существенных изменений актуального разнообразия, хотя возможны и другие варианты. В первом случае графики типа 20, а, в для меры морфологического разнообразия, построенные с учетом реаль-ной истории изучения, должны быть асимптотическими.
Полученный при полном исследовании график N2/N1 с выраженной горизонтальной правой частью дает обнадеживающие указания на возмож-но достаточную полноту данных, не доказывая ее, но делая более вероят-ной, и позволяет далее детализировать исследования до уровня видов и в перспективе - меры морфологического разнообразия. Для приемлемой точности отношения N2/N1 достаточно, как указывалось выше, чтобы изу-ченности dt на сравниваемых границах не слишком отличались. При хара-ктеристике процесса в целом необходимы также сведения о появлении и вымирании, которые тоже могут контролироваться с помощью графиков отношений DN+/N1 и других такого типа. Для появлений и вымираний тре-бования к материалу более жесткие, так как в этом случае главную роль играет полнота изученности dt, и риск получения неверных результатов возрастает. Ниже будет указан другой способ оценки приращений DN+ и DN-.
Асимптотический метод кажется привлекательным своей свободой от каких-либо сложных предварительных предположений, единственная посылка: актуальное разнообразие есть постепенно увеличивающаяся в хо-де исследований часть потенциального, но простота исходного положения также и ограничивает возможности этого подхода.
Экстраполяционный “метод”
Экстраполяционный подход к оценке полноты палеонтологического материала может быть охарактеризован только на уровне общей идеи и не-которых предварительных соображений; он еще отсутствует как рабочий метод и даже его осуществимость нуждается в обосновании. Обсуждение лишь гипотетической возможности кажется оправданным в связи с высо-кой перспективностью экстраполяционного подхода в случае удачной ре-ализации.
Идея подхода заимствована из современных фаунистических иссле-дований, где соответствующий метод применяется для оценки полного чи-сла видов в локальных фаунах (Песенко, 1982). Распределение частот ви-дов по числу экземпляров в больших коллекциях, согласно многочислен-ным исследованиям (см. цит. работу Песенко), подчиняется логарифми-чески нормальному (логнормальному) распределению. Эмпирическое рас-пределение усеченное, левая часть полной кривой распределения в нем не представлена. При формальной интерпретации площадь под полной кри-вой нормального распределения логарифмов численностей соответствует числу видов в исследуемой фауне, а ее отношение к площади усеченного эмпирического распределения легко вычисляется. Прогноз общего числа видов подразумевает неизменную методику сборов, в палеонтологии так-же выделения материала из породы и стабильность практических критери-ев ранга таксонов.
Таким образом, описываемый подход предполагает экстраполяцию имеющихся данных до числа таксонов, соответствующего dt=1, для ло-кальных или региональных ископаемых фаун на коротких временных ин-тервалах с последующим расширением их до полного глобального разно-образия (см. следующий раздел). Этим он отличается от примененного Ф. Сигнором (Signor, 1985) непосредственного приложения логнормально-го распределения сразу к глобальному материалу по периодам.
Оценка общего числа таксонов, существовавших в некотором вре-менном интервале, является основной для определения мгновеных значе-ний и числа появившихся и вымерших таксонов, причем для всех величин d=1 (то же относится к работе с мерой морфологического разнообразия). Наиболее надежным представляется получение необходимых величин из очевидных равенств:
1) число таксонов N(t1/2) на границе двух последовательных временных интервалов t1 и t2
N(t1/2) = N(t1) + N(t2) - N(t1+t2),
N(t1), N(t2) - число таксонов в первом и втором интервалах, N(t1+t2) – чис-ло таксонов в обоих интервалах вместе;
2) число появившихся таксонов
DN+ = N(t) - N(t1),
N(t) - общее число таксонов в интервале t, N(t1) - число таксонов на его нижней границе;
3) число вымерших таксонов
DN- = N(t) - N(t2),
N(t2) - число таксонов на верхней границе интервала.
Очевидны серьезные преимущества этого способа оценки и компен-сации неполной палеонтологической изученности. Снимается большинст-во связанных с ней вопросов, в том числе относящихся к филогенетичес-ким связям, недоучет которых при неполной изученности может повлечь за собой неверную фиксацию моментов появления и вымирания таксонов (Patterson, Smith, 1989). В результате экстраполяции до d=1 сложности, за-висящие от неточной филогении, в значительной мере сглаживаются (но сохраняется погрешность, зависящая от филетического вымирания). Появ-ляется возможность оценить погрешности всех получаемых величин и отобрать для работы пригодные данные на подходящем таксономическом уровне или на уровне меры. Совместно с детальностью хронологической шкалы, задаваемой автоматически в зависимости от состояния материала, это обеспечивает контроль качества исходных данных и получаемых вы-водов на всех этапах работы, что является тривиальным условием совре-менного научного исследования.
Подход назван здесь экстраполяционным, чтобы отразить общую идею, не связывая ее с конкретным видом распределения. Необходимость экстраполяции определяет жесткие требования к используемым функци-ям: они должны быть хорошо обоснованы теоретически. К сожалению, этого пока нет, но обоснование выбора адекватных функций, если оно ока-жется возможным, несомненно оправдает затраченные усилия. В совре-менной фаунистике изучаются мгновенные распределения, тогда как иско-паемый материал относится к временным интервалам, что может повлиять на вид распределения, в том числе оно может меняться с увеличением про-должительности интервалов.
Палеонтологические данные почти не изучены эмпирически. Очень важные исследования проведены Кохом (Koch, 1987; Koch, Morgan, 1988) на материале меловых и голоценовых моллюсков Северной Америки. Он столкнулся с трудностями при попытках выяснить число экземпляров в старых коллекциях, но успешно продемонстрировал возможность замены его числом коллекций, что представляет большой интерес. Кох использо-вал логарифмический ряд Фишера, не позволяющий экстраполировать до dt=1, но в его совместной публикации с Морганом отмечена принципиаль-ная возможность работы с логнормальным распределением, которое при-менялось им в более ранних работах (Koch, 1978). Оно же было использо-вано в некоторых исследованиях по эволюции видового разнообразия (Sepkoski, 1993). По Песенко (1982), в современной фаунистике логариф-мический ряд Фишера и логнормальное распределение равноправны, при-чем выбор предпочтительного не удается обосновать. Исследования на па-леонтологическом материале являются весьма актуальными.
Более подробное обсуждение экстраполяционного подхода приме-нительно к ископаемому материалу кажется преждевременным. В случае доказательства его невозможности в описанном варианте, что очень веро-ятно, может быть применен простой метод, иллюстрируемый рис. 20, ж. Хотя для однозначной интерпретации данного рисунка, как указано выше, требуются некоторые дополнительные сведения, он наглядно показывает вид кривой, которая может характеризовать состояние высокой палеонто-логической изученности. Как и в асимптотическом методе, принимается фиксированная стратиграфическая корреляция и, кроме того, состав таксо-нов в соответствии с современными представлениями. Почти так построе-ны левые кривые на рис. 20, отличаясь тем, что обсуждаемые здесь графи-ки строятся для временных интервалов, а не границ. По оси абсцисс долж-но быть число экземпляров в коллекциях, накопленных до определенных дат, по оси ординат - число таксонов или мера морфологического разнооб-разия в соответствии с теми данными о распространении, которые следу-ют из стратиграфического положения экземпляров, учтенных по оси абс-цисс (исторический аспект, игнорируемый на рис. 20). Мера разнообразия предпочтительнее, когда на графике для числа таксонов вырисовывается его постоянное увеличение, как в разобранном выше случае кампанского разнообразия гастропод. Если применение меры невозможно в связи с не-подготовленностью материала, то придется отказаться от исследования приращений DN+, DN- и ограничиться изучением отношения N2/N1 и срав-нительным анализом роли таксономических ревизий и нового коллекцион-ного материала.
Перспективным может быть построение графиков для увеличиваю-щихся временных интервалов, так как изученность может зависеть от их продолжительностей. Значительная часть исходного материала содержит-ся в обычных палеонтологических видовых картотеках и компьютерных базах данных. Выяснение числа экземпляров может оказаться затрудни-тельным, эти осложнения могут преодолеваться заменой числа экземпля-ров на другую подходящую переменную, связанную с ним монотонной за-висимостью, но интерпретация графиков может осложниться. Неприятной особенностью метода (как и асимптотического) может стать отсутствие асимптотической части в слишком многих случаях и вытекающая из этого необходимость дальнейших сборов и неопределенно долгого ожидания, пока асимптота проявится. Не исключено стереотипное строение графи-ков, допускающее экстраполяцию до асимптоты при неполной, но доста-точно высокой изученности.
В морских условиях площадь осадконакопления в общем совпадает с площадью обитания организмов. С некоторым огрублением потенциаль-ное разнообразие морских организмов с твердым скелетом можно считать соответствующим их полному прижизненному разнообразию в районах сборов палеонтологических коллекций. Если выявлено потенциальное раз-нообразие, вопрос о полном древнем разнообразии, региональном или гло-бальном, сводится к двум другим. Во-первых, какое дополнительное раз-нообразие может быть связано с закрытыми территориями, и во-вторых, какое разнообразие невосполнимо утеряно в результате постседиментаци-онного разрушения древних отложений.
Для континентальных фаун и флор может быть велика роль организ-мов, обитавших в зонах отсутствия осадконакопления и плохо сохраняю-щихся. Это определяет сложность и меньшую достоверность реконструк-ции их разнообразия и ряд ее особенностей. Ниже подразумеваются преи-мущественно морские организмы.
Площадь отложений, содержащих потенциальное разнообразие, со-ответствует части площади обитания представленных в этих отложениях древних сообществ организмов. За счет чередования разнофациальных от-ложений один разрез может характеризовать древнюю фауну или флору значительных районов, площадь которых в общем случае неизвестна. Вы-страивается последовательность: актуальное разнообразие, полученное с определенных территорий, - потенциальное разнообразие, характеризую-щее какие-то неустановленные площади прошлого распространения древ-них фаун и флор, - полное разнообразие, относящееся ко всей области их былого существования. Последнее может оцениваться с помощью графи-ков площадь - разнообразие путем экстраполяции на всю древнюю пло-щадь обитания.
Количественные исследования зависимости разнообразие - площадь на ископаемом материале очень немногочисленны и ориентированы нес-колько иначе (Schopf, 1974; Sepkoski, 1976; Peters, Foote, 2001). Здесь мож-но только кратко отметить несколько вопросов.
Зависимость разнообразие - площадь подробно рассмотрена в совре-менной биогеографии для островов и неизолированных территорий. Ре-зультаты не вполне однозначны (Песенко, 1982), но размеры изучавшихся площадей, как правило, меньше имеющих значение для обсуждаемой про-блемы древнего разнообразия. Важным является заключение Е. Коннора и Е. Маккоя (Connor, MacCoy, 1979) о сигмоидной форме кривых площадь - число видов при распространении исследований на большие территории. По Ю. Г. Пузаченко и Н. Н. Дроздовой (1986), для больших континенталь-ных территорий число видов растет медленнее логарифма площади. В ука-занной работе теоретически исследовалась площадь охраняемых террито-рий, необходимая для сохранения биологического разнообразия. Она же есть площадь, достаточная для выявления полного состава фауны или фло-ры. Для европейской России желательная охраняемая площадь - 1% всей ее площади, а в гетерогенных условиях Горного Алтая - 4,5%. Предпола-гается специальный выбор положения охраняемых территорий, что не со-ответствует условиям сбора ископаемых организмов, но может хотя бы ча-стично компенсироваться фациальным чередованием отложений. В морс-кой биогеографии продемонстрировано быстрое, по показательному зако-ну, уменьшение приростов числа элементарных биогеографических выде-лов при увеличении числа используемых для районирования видов (Ней-ман, Зезина, Семенов, 1977). Имеются все основания считать зависимость площадь - разнообразие асимптотической или очень близкой к асимптоти-ческой. Эмпирические исследования палеонтологического материала представляют большой интерес.
При построении кривой площадь - разнообразие с целью выявления полного разнообразия нельзя использовать актуальное разнообразие, так как тогда не обеспечивается выполнение условия d=1, следовательно орди-ната - потенциальное разнообразие, а абцисса - площадь его обитания. В случае асимптотического поведения кривой для очень крупных регионов или глобальной вопрос о реальной площади обитания потенциального раз-нообразия не имеет большого значения. Теоретически она может оцени-ваться как площадь соответствующих биохорий на палеобиогеографичес-ких реконструкциях. Это придало бы всему построению весьма спекуля-тивный характер, но асимптота может выявиться и при подходящей упро-щающей замене независимой переменной даже при искажении ее соответ-ствия площади биохорий. Возможность огрубляющей замены сильно зави-сит от того, какую часть полной площади обитания занимает асимптоти-ческая часть кривой разнообразие - площадь.
Закрытые территории могут служить полигоном для выяснения ре-гиональной полноты фаун, изучаемых преимущественно по сборам из об-нажений, и надежности прогноза полного регионального разнообразия. Такую возможность дают микропалеонтологическое изучение кернов и сборы в шахтах, а особенно - в глубоких открытых карьерах, когда они находятся на большом расстоянии от естественных обнажений в перифе-рийных частях осадочных бассейнов. Большой интерес в этом отношении представляет также возможность использовать подходы, предлагаемые секвентной стратиграфией (Holland, 2001). Изученность комплексов иско-паемых организмов может быть высокой на фоне неполной изученности отдельных точек сбора, вследствие суммативности местных и региональ-ных фаун и флор.
Разрушение отложений различными геологическими процессами мо-жет иметь большое значение, когда полностью или почти полностью исче-зают древние биогеографические провинции. Тогда при глобальном ана-лизе возможны попытки ввести поправки (с учетом градиентов разнообра-зия). Задача может быть легче, если провинция хотя бы фрагментарно из-вестна по отложениям смежных возрастов. Субдукция океанической коры, уничтожающая громадные площади древних отложений, не имеет почти никакого значения, так как число глубоководных видов по разным оцен-кам в 30-70 раз меньше числа шельфовых и верхнебатиальных видов (Ви-ноградова, 1977), но отдельная оценка желательна для вклада океаничес-ких островов. Преобладающее значение мелководного разнообразия дает возможность упростить выбор независимой переменной для кривых пло-щадь – разнообразие. Может быть достаточным грубый подсчет площади эпиконтинентальных морей вместе с произведением характерной ширины шельфов на их длину.
Захороненное разнообразие уничтожается преимущественно круп-ными блоками в связи с разрушением или метаморфизацией крупных масс вмещающих отложений. Это вызывает определенные сомнения в адекват-ности описания потерь разнообразия как пуассоновского процесса (Pease, 1985).
Асимптотическое или близкое к нему поведение кривой площадь - разнообразие не противоречит известным представлениям о влиянии на разнообразие трансгрессивно-регрессивных циклов, но позволяет уточ-нить их. Если разнообразие приближается к полному на площадях, сущес-твенно меньших всей площади обитания, даже значительные колебания уровня моря могут никак не сказываться на нем. Например, не выявлено сколько-нибудь заметного избыточного вымирания родов и семейств в связи с гляциоэвстатической регрессией в антропогене, хотя имеются ре-конструкции, описывающие кратковременное снижение числа видов (Wise, Schopf, 1981). Отсутствие вымирания надвидовых таксонов при антропогеновой регрессии благоприятно для оценки перспективности кри-вых площадь - разнообразие в изучении полного древнего разнообразия по крайней мере на уровне выше вида. Для регрессивных фаз характерна меньшая доступность ископаемых остатков из нормально-морских отло-жений и поэтому меньшая изученность фауны. Это может привести к по-явлению ложных минимумов на кривой разнообразия и обусловить кол-лективное поведение групп, описанное в разделе о второй интерпретации кривых.
Очевидно, при выявлении полного разнообразия древних фаун и флор должна быть исследована достаточно большая часть древней площа-ди обитания при более или менее равномерной высокой изученности раз-ных территорий. В континентальных отложениях выполнение этих усло-вий проблематично, хотя положение может облегчаться аллохтонностью многих захоронений со смешением остатков из разных обстановок обита-ния. Во всяком случае, сама попытка построения кривых площадь – разно-образие предоставляет возможность контролировать как исходные данные в отношении полноты охвата территорий, так и зависящие от этого резуль-таты. При существенной неравномерности данных по территориям велика опасность принять региональные события, зависящие от многих причин, как резкие смещения климатических и биогеографических границ, смены ландшафтов, наличие перерывов в осадконакоплении, в том числе нерас-познанных, и т. д., за крупные глобальные изменения разнообразия.
Заключение
Для перехода от кривых числа таксонов, которые всегда могут быть построены независимо от их адекватности изменениям разнообразия орга-низмов прошлого, к кривым разнообразия в точном смысле необходим ко-мплекс исследований различного характера. Современные кривые числа таксонов не подразумевают контроля качества результатов и поэтому не могут быть отнесены к области проверенного знания. Реалистичные кри-вые должны в большей степени зависеть от состояния знаний: на каких-то участках может быть отсутствовать вообще, где-то прорисовываться толь-ко пунктиром. Хотя общий ход изменений фанерозойского разнообразия во многом достаточно установлен (но не во всем, см. “Pull of Recent” в раз-деле о мере), задача состоит в повышении разрешающей способности. Лю-бые незначительные изменения разнообразия, в частности малые миниму-мы, не могут быть надежно выявлены и охарактеризованы без осуществле-ния всего необходимого комплекса исследований, в котором вопросы обо-снованности результатов приобретают решающее значение.
Прямое прочтение изменений актуального разнообразия как эволю-ции полного разнообразия некорректно. Оно не может быть оправдано распространенными представлениями о постепенном приближении сегод-няшних реконструкций процесса к его все более адекватному описанию по мере дальнейшего изучения. Актуальное разнообразие при накоплении но-вого материала и перехода на более низкие таксономические уровни в дей-ствительности все более точно характеризует только потенциальное разно-образие. Последнее не обязательно репрезентативно по отношению к пол-ному разнообразию (пониженная изученность нормально-морской фауны в регрессивные фазы и др.), поэтому воспроизводимость результатов в данном случае не может служить обоснованием их адекватности.
Выше описан ряд процедур, позволяющих сделать изучение древне-го разнообразия более рациональным. Кажется необходимым обеспечить возможность обоснованного алгоритмизированного выбора пригодных для изучения групп организмов, уровня детальности исследований разно-образия (ранга таксонов или меры морфологического разнообразия) и дро-бности хронологической шкалы, а также контроля полноты данных по тер-риториям. Результатом должна быть оптимизация процесса изучения в рамках реальных возможностей конкретного палеонтологического матери-ала: при изучении числа таксонов все более высокого ранга уменьшение погрешностей, зависящих от изученности, сопровождается потерей точно-сти описания морфологического разнообразия.
В трехчленной схеме перехода от актуального разнообразия к потен-циальному и далее полному отчетливо нарастает сослагательность изложе-ния. Это связано с неразработанностью двух последних ее частей, форму-лируемых здесь только как идеи о возможных направлениях поиска. При попытках реализации схемы гипотетическая составляющая должна нарас-тать в той же последовательности вследствие объективных особенностей материала.
Ряд рассмотренных выше разнообразий (актуальное - потенциальное - полное) является незавершенным. Биотические отношения, влияющие на дивергенцию и вымирание, могут устанавливаться в группах таксонов, часть которых не сохраняется в палеонтологической летописи. Отсюда следует необходимость дополнения ряда разнообразий замыкающим чле-ном - общим разнообразием, включающем, кроме хорошо фоссилизирую-щихся групп, также и всех мягкотелых. Вопрос об эволюции общего раз-нообразия находится преимущественно за пределами непосредственного анализа эмпирического материала, отображаемого кривыми разнообразия, и может частично решаться лишь на основе косвенных гипотетических со-ображений.
Анализ диверсификации древних организмов включает исследова-ние роли нескольких взаимовлияющих переменных, обусловленных состо-янием изученности крупных групп: полноты актуального разнообразия по отношению к потенциальному, изменений потенциального разнообразия в результате прогресса морфологии скелетных остатков, членения разно-образия на таксоны. Тем самым включается механизм научной саморе-флексии - характеристики развитого знания, вплоть до исследования осо-бенностей принятия таксономических решений, определяемых объектив-ными психологическими закономерностями распознавания и классифици-рования. Значение внешнего по отношению к палеонтологии (в узком смысле) фактора - стратиграфической корреляции - также может быть изу-чено. Очевидной целью анализа является выявление количественных хара-ктеристик, асимптотически постоянных в истории изучения групп. Прин-цип изучения общеизвестен: фиксация переменных, кроме изучаемой, но в палеонтологии он пока-что почти не применяется.
Программа исследований может осуществляться только при участии широкого круга специалистов. Дж. Сепкоски выполнил громадную работу по сведению данных о распространении семейств и родов, заложив основы дальнейшего продвижения, но это уже близко к пределу индивидуальных возможностей. Новые обобщения будут возможны, если исследование из-менений разнообразия станет рутинной частью обычных палеонтологичес-ких работ по группам организмов. Необходима некоторая унификация способов представления исходных данных, допускающая их использова-ние при пересмотре стратиграфической корреляции или хронологической шкалы.
Несомненно большое биологическое значение правильной интерпре-тации кривых разнообразия. Определенную роль могут сыграть математи-ческие модели, но здесь хотелось бы обратить внимание на менее очевид-ную сторону проблемы. Стереотипный ход кривых для числа таксонов разного ранга, их простая связь между собой и часто простые зависимости N+(t), N-(t) на длительных временных интервалах свидетельствуют о воз-можности феноменологического описания типичных вариантов диверси-фикации. Отсюда следует принципиальная возможность поставить обрат-ную задачу - определение продолжительностей геохронологических под-разделений (Дмитриев, 1978; Odin, Odin, 1990). Ее решение выходит за рамки интересов только палеонтологии и биологии и представляется весь-ма актуальным, учитывая плохое состояние изотопно-геохронологической шкалы (Рублев, 1996). Интерполяция возраста границ между точками опорных изотопных датировок по гладким кривым уже в настоящее время может существенно сузить разброс продолжительностей веков, свойствен-ный современным шкалам. Полное выяснение этого вопроса может дать многие и неожиданные результаты.
(в соавторстве с А.Г.Пономаренко)
Изучение эволюции таксономического разнообразия континенталь-ных экосистем представляет собой сложную и рискованную задачу в связи с значительной фрагментарностью неморской палеонтологической летопи-си. Это не должно быть основанием для отказа от попытки исследования, так как оценить предполагаемое влияние возникающих осложнений можно только в результате анализа картины диверсификации, полученной при непосредственном изучении актуального разнообразия континенталь-ной биоты.
Диверсификация неморских организмов изучена явно недостаточно. Для морских организмов имеются многочисленные обзоры и конкретные исследования, которые в совокупности дают общее представления о про-цессе. В отличие от них континентальное биоразнообразие обсуждается только в единичных работах. По водным и амфибиотическом животным еще недавно имелся лишь обзор на уровне отрядов, выполненный Дж. Грей (Gray, 1988), на основе которого были произведены некоторые вторичные исследования (Дмитриев и др., 1994а,б). Позднее были сообще-ны первые результаты изучения диверсификации пресноводных животных на уровне семейств (Дмитриев и др., 1995), основанного на оригинальном материале, собранном сотрудниками Палеонтологического института РАН.
Сведения о семействах насекомых одновременно и независимо обоб-щены в Англии (Jarzembowski, Ross, 1993, Ross, Jarzembowski, 1993) и в США (Labаndeirа, Sepkoski, 1993). Как отмечалось в откликах на послед-нюю работу (Doyle, 1993), ее результаты не вполне достоверны. Англий-ское исследование выполнено на основе датировок появления и вымира-ния семейств с детальностью лишь до эпохи и не выявляет кратковремен-ных событий. Сравнение описанных ниже результатов с полученными и в том и в другом исследовании затруднено использованием в обоих тради-ционного построения кривых разнообразия.
По тетраподам имеются исследования М. Бентона (Benton, 1985, 1987, 1989), а также Н.Н. Каландадзе и А.С. Раутиана (1993) на уровне се-мейств. В обоих использовано традиционное построение кривых, в первом с детализацией датировок появления и вымирания до веков, во втором - до эпох.
Семейства сосудистых растений и отдельно покрытосемянных рас-сматривались М. Бентоном (Benton, 1993) традиционным способом по эпо-хам. К сожалению, не удалось получить материала по растениям для неза-висимого анализа в данной работе, хотя знание их диверсификация несом-ненно чрезвычайно важно для понимания эволюции континентального, биоразнообразия и происходивших при этом процессов.
В настоящем исследовании обобщены современные материалы по изменениям актуального разнообразия пресноводных животных, насеко-мых и тетрапод на уровне семейств с детализацией временной шкалы до веков. В заключении к данной главе сделаны выводы о тех характеристи-ках процесса, которые могут быть достаточно надежно установлены без специального анализа полноты актуального разнообразия.
Водные животные
Изучались изменения числа семейств животных - обитателей вну-триконтинентальных водоемов в течение всей истории их существования. Использованы сведения о возрастных диапазонах распространения 915 се-мейств, подготовленные преимущественно сотрудниками Палеонтологи-ческого института РАН. Группы, охарактеризованные лишь редкими и разрозненными находками (простейшие, черви, губки, мшанки; всего око-ло 20 семейств) не учитывались. Данные по моллюскам (97 семейств) пре-доставлены Я.И. Старобогатовым (ЗИН), время появления и вымирания почти всех семейств указано с точностью только до эпохи. Членистоногие представлены 304 семействами (из них насекомых - 174), данные собраны коллективом Лаборатории артропод: В.А. Благодеровым, К.Ю. Еськовым, Е.Д. Лукашевич, В.Г. Новокшеновым, А.Г. Пономаренко, Ю.А. Поповым, Л.Н. Притыкиной, Н.Д. Синиченковой, И.Д. Сукачевой. Список семейств членистоногих (кроме насекомых) был просмотрен А.С. Алексеевым (МГУ), сделавшим некоторые уточнения. Сведения о костистых рыбах со-браны А.Г. Пономаренко и откорректированы А.Ф. Банниковым, осталь-ные рыбы и бесчелюстные даны О.А. Лебедевым; всего учтено 272 семей-ства бесчелюстных и рыб. Диапазоны распространения 58 семейств пре-сноводных и околоводных птиц получены от Е.Н. Курочкина, 184 се-мейств остальных тетрапод - от Н.Н. Каландадзе и А.С. Раутиана.
Результаты обработки перечисленных данных изображены на рис. 21-26, кроме того, для сравнения на рис. 21 приведены кривая для се-мейств морских животных по данным Сепкоски (Sepkoski, 1982, с после-дующими дополнениями) и кривая по 179 отрядам пресноводных живот-ных.
До изложения конкретных результатов следует сделать несколько за-мечаний.
Животные - обитатели внутриконтинентальных водоемов иногда для простоты изложения ниже называются пресноводными, хотя некоторые из этих водоемов могли быть с повышенной соленостью.
<![if !vml]> <![endif]>
<![endif]>
Рис. 21. Изменения разнообразия животных в морской и внутриконтиненталь-ной водной биотах в координатах (t,N) (Дмитриев и др., 1995): а - морская биота, се-мейства; б, в - неморская биота; б - семейства, в - отряды
Как водные ниже рассматриваются животные, полностью обитаю-щие в воде на всех стадиях жизненного цикла, и амфибиотические группы (многие насекомые, водоплавающие птицы и др.).
До карбона диагностика неморских условий обитания часто затруд-
нительна. Иногда она трудна и для более молодых отложений.
Эволюция водных животных на континентах не автохтонна. Часть внутриконтинентальной водной биоты формируется за счет вселения как из моря, так и с суши. Становление биоты происходило в результате миг-рации морских животных, роль которых была определяющей на ранних этапах ее развития.
Изменения числа отрядов и семейств пресноводных животных (pиc. 21) рaccматривались в cpaвнeнии c динaмикoй paзнooбpaзия мopcкиx фa-yн. Число отрядов (рис. 21, в) пресноводных животных изменяется в целом сходно с изменениями числа семейств (рис. 21, б), особенно наглядно сов-падение или близкое соответствие основных минимумов разнообразия. Как у отрядов, так и у семейств в пaлeoзoйcкoй чacти графика oтcyтcтвуют пеpиoды cтaбилизaции paзнooбpaзия. Рaзнooбpaзиe пресноводных живот-ных после ордовика в целом pacтeт, a зaтeм в пoзднeм кapбoнe и пepми снижается. В море в палеозое длительно поддерживается динамическое равновесие разнообразия на уровне около 400 семейств (рис. 21, а). На кривой разнообразия отрядов обращает на себя внимание относительно более быстрый рост числа отрядов по сравнению с семействами (девонс-кий максимум составляет 35% числа современных отрядов, известных в ископаемом состоянии, и лишь 13% семейств, карбоновый 50 и 25% соот-ветственно). Сходные соотношения характерны и для морских животных (карадокский максимум: отрядов - 79%, семейств - 42%). В мезозое и кай-нозое разнообразие пресноводных животных преимущественно возрастает до современности, как и морских.
На кривой отрядов (рис. 21, в) более или менее обособлена только часть до конца девона, затем в целом наблюдается достаточно стабильное увеличение разнообразия, лишь осложняющееся сравнительно слабо выра-женными эпизодами его снижения. Сходная картина для отрядов была описана ранее по данным Дж. Грей (Gray, 1988; Дмитриев и др., 1994а,б), но сведения по семействам тогда отсутствовали. Теперь имеется возмож-ность соотнести результаты по отрядам и семействам. Так как отрядов в пять раз меньше, чем семейств, кривая для них оказывается более обоб-щенной.
Ордовик. Первое зарегистрированное в палеонтологической летопи-си вхождение животных в биоту внутриконтинентальных водоемов, по данным Г. Греффина (Graffin, 1992), относится к аренигу, когда появились пресноводные представители бесчелюстных семейства Arandaspididae. В лландейло к ним добавились представители семейств Astraspididae и Eri-ptichiidae. Только последние из них переходят в силурийскую внутрикон-тинентальную водную биоту, в связи с чем число семейств снижается в позднем ордовике с двух до одного, и появляется слабо выраженный ми-нимум на границе ордовика и силура. Он соответствует минимуму в разно-образии морских животных, но материал по неморским слишком скуден.
Силур. В силуре общее разнообразие семейств формируется позво-ночными (почти целиком бесчелюстными) и членистоногими (почти цели-ком хелицеровыми), вклад которых примерно одинаков. Обе крупные группы в течение почти всего периода увеличивают свое разнообразие, од-нако в конце его ведут себя по-разному. Разнообразие членистоногих про-должает увеличиваться до границы с девоном (но не удалось получить от-дельную точку для границы лудлова и пржилола), тогда как у позвоноч-ных оно несколько уменьшается в пржидоле (рис. 22, а, б). Суммарная кривая (рис. 21, б) на пржидольском участке становится более пологой и характеризует замедление роста общего разнообразия, отличаясь от кри-вой для семейств морских животных (рис. 21, а), на которой границе с де-воном соответствует отчетливый, хотя и не глубокий, минимум.
Девон. Хотя в девоне появляются первые представители неморских двустворчатых моллюсков, а в самом конце его первые тетраподы, они практически не влияют на суммарную кривую разнообразия семейств, ход которой по-прежнему зависит от изменений разнообразия бесчелюстных, рыб и членистоногих. У последних в девоне происходит медленное увели-
<![if !vml]> <![endif]>
<![endif]>
Рис. 22. Изменения числа семейств в основных крупных группах неморских вод-ных животных в координатах (t,N) (Дмитриев и др., 1995): а - бесчелюстные и рыбы, б - членистоногие (кроме насекомых). в - моллюски. г - тетраподы (кроме птиц), д – на-секомые, е - птицы
чение числа семейств, на кривой (рис. 22, б) это отражается в виде почти прямого субгоризонтального участка. Два минимума на общей кривой раз-
нообразия - в конце раннего девона и на границе с карбоном (рис. 21, б) определяются развитием бесчелюстных и рыб. Первые почти целиком вы-мирают в зигене и эмсе (34 семейства, снижение разнообразия на 21 се-мейство), разнообразие рыб в зигене увеличивается на 10 семейств (появ-ляется 12), тогда как в эмсе только на 2 (появляется 8). Таким образом, в зигене можно предположить конкурентное вытеснение бесчелюстных ры-бами без существенного изменения общего разнообразия. В эмсе вымира-ние рыб более выражено и нет существенного роста их разнообразия. В морской биоте аналога раннедевонского вымирания пресноводных позво-ночных не выявлено. Минимум на границе девона и карбона на неморской кривой полностью определяется уменьшением числа семейств рыб.
Карбон. В составе фауны внутриконтинентальных водоемов появля-ются первые насекомые (московский век), их разнообразие почти до сере-дины перми продолжает оставаться стабильно низким (рис. 22, д). Также низким разнообразием семейств характеризуются каменноугольные мол-люски (рис. 22, в). Основной вклад в изменения общего разнообразия вно-сят рыбы, тетраподы и членистоногие (без насекомых, так же и далее). Чи-сло семейств рыб в турне увеличивается, от визе до позднего карбона поч-ти не меняется, в конце карбона несколько снижается. Разнообразие тетра-под и членистоногих в течение карбона возрастает, у тетрапод - до конца периода, у членистоногих - до конца среднего карбона, а в позднем умень-шается. На границе карбона и перми имеется неярко выраженный мини-мум общего разнообразия (отчетливое снижение в позднем карбоне, но очень слабый рост разнообразия в ассельском веке). Минимум соответст-вует таковому на кривой разнообразия морских животных. В отличие от него небольшое снижение разнообразия морских животных в конце ранне-го карбона не имеет аналога в неморских водоемах, но в исходном матери-але не удалось разделить серпуховский и башкирский века, которые здесь рассматриваются совместно.
Пермь. На кривой суммарного разнообразия неморских водных жи-вотных пермь выделяется постепенным снижением числа семейств, что в целом хорошо согласуется с продолжительным снижением разнообразия морских животных. В деталях ход кривых несколько различается в их кун-гурско-казанской части. Постепенное снижение разнообразия хорошо вы-ражено (рис. 22) в развитии членистоногих и тетрапод, несколько менее ярко у рыб, слабо среди моллюсков. Из пяти крупных групп несколько увеличивают разнообразие лишь водные насекомые, но их история близ границы с триасом изучена недостаточно.
Триас. Триасовое развитие характеризуется четкими минимумами в начале и конце периода, совпадающими с минимумами морского разнооб-разия. Максимум между ними у неморских животных смещен ближе к концу периода. Снижение разнообразия в противоположность длительно-му пермскому сосредоточено полностью в рэте, тогда как морское, как и в перми, растянуто во времени (о родах см. главу о кривых разнообразия). Уменьшение общего разнообразия в позднем триасе определяется умень-шением числа семейств рыб, тетрапод и членистоногих, тогда как разнооб-разие моллюсков и водных насекомых в позднем триасе несколько увели-чивается. В раннем и среднем триасе диверсификация всех изученных групп происходит согласованно, только у насекомых она выражена менее отчетливо.
Юра. Разнообразие монотонно увеличивается в течение всего перио-да, в средней юре медленнее, чем в ранней и особенно в поздней. В море разнообразие также постоянно нарастает, но более равномерно. При ана-лизе изменений числа семейств в крупных группах (рис. 22) обращает на себя внимание сохранение в ранней и средней юре стабильно низкого раз-нообразия рыб, тетрапод, членистоногих и моллюсков. У моллюсков оно даже несколько снижается по сравнению с концом триаса (рис. 22, в). У рыб и тетрапод ранне-среднеюрский уровень разнообразия составляет око-ло 1/3 пермского максимума. Для этих групп характерна также низкая ско-рость появления семейств. Скорость появления у рыб и тетрапод в ранней и средней юре составляет 0.15 семейства/млн. лет, тогда как в поздней юре семейства тетрапод появлялись со скоростью 0.60, как и рыб в триасе (0.62). Соответствующие удельные скорости составляют соответственно 0.022-0.024 против 0.058-0.065 1/млн. лет. Эта длительная эпоха понижен-ных наблюдаемых скоростей является очень необычной. Раннеюрский рост общего разнообразия пресноводных животных определяется в основ-ном диверсификацией водных насекомых. В поздней юре происходит бо-лее активный рост разнообразия также водных тетрапод, в меньшей мере – моллюсков и членистоногих.
Ранний мел. Раннемеловое развитие характеризуется продолжаю-щимся увеличением разнообразия неморских водных животных, включая появившихся в раннем мелу водных птиц. Значительно увеличивается чис-ло семейств водных и амфибиотических четвероногих, в остальных груп-пах не наблюдается выраженного роста, рыбы продолжают вялое разви-тие, характерное для юры. Отличие от диверсификации морских живот-ных проявляется только в конце эпохи, когда в альбе число неморских се-мейств незначительно снижается (рис. 21, б). В морской биоте аналог это-го снижения разнообразия не выявляется. На рис. 22 апт и альб не разделе-ны на кривых для рыб, членистоногих и тетрапод. Данные объединены для более полного учета семейств, находки которых сделаны в отложениях, датируемых не точнее, чем апт или альб. При этом некоторые различия по-ведения групп по векам на графике оказываются замаскированными, что заставляет описать их в тексте.
Апт. Некоторое снижение числа семейств наблюдается только у рыб. На рис. 21 оно скомпенсировано преобладающим появлением новых семейств насекомых и других членистоногих и тетрапод.
Альб. У рыб и водных насекомых вымирания или появления новых се-мейств не установлено. По данным Н.С. Калугиной (1980), в позднем ме-лу, в отличие от раннего, очень редко захоронялись насекомые - обитатели стоячих водоемов; их разнообразие, возможно, уменьшается в конце ран-него мела на более низких таксономических уровнях. Среди птиц появля-ется одно новое семейство и еще одно у других тетрапод. Незначительное (на одно семейство) снижение разнообразия на рис. 21 определяется пос-ледними находками в альбе трех семейств тетрапод.
Поздний мел. В позднем мелу происходит согласованное увеличе-ние наблюдаемого разнообразия всех групп. Кривые разнообразия морс-ких и неморских водных животных для этого времени весьма сходны, од-нако маастрихтское вымирание в пресных водах, в отличие от морской биоты, удается выявить не в большинстве групп. Минимум суммарной кривой на границе с данием связан с уменьшением числа семейств в ма-астрихте только у тетрапод, возможно, включая птиц. Минимум разнооб-разия обеих групп показан на верхней границе дания, но для птиц это мо-жет быть связано с неполнотой данных, из-за чего нет точки на границе дания и маастрихта. У прочих тетрапод минимум смещен на границу па-леоцена в связи с предполагаемым датским возрастом последних находок динозавров. Данные по моллюскам, рыбам и членистоногим (в том числе и насекомым) для конца мела недостаточно детальны. У насекомых не изве-стны представительные местонахождения в маастрихте. Пaдeниe paзнooб-paзия семейств пресноводных организмов в кoнцe мeлa сравнительно с на-блюдаемым в морской биоте cлaбoe.
Кайнозой. В целом в кайнозое разнообразие возрастает и в пресно-водной и в морской биоте, лишь в среднем и позднем миоцене на кривой для неморских животных имеется субгоризонтальный участок. Резкое уве-личение их разнообразия в позднем эоцене - несомненный артефакт, свя-занный с очень большим кажущимся появлением новых семейств насеко-мых в балтийском янтаре. Это определяется громадными коллекциями на-секомых из балтийского янтаря, несопоставимыми с какими-либо другими коллекциями. На кривой для насекомых (рис. 22, д) позднему эоцену соот-ветствует четкая ступень. Выраженный подъем разнообразия в олигоцене можно видеть на кривых для рыб, птиц и наземных тетрапод. Если исклю-чить позднеэоценовую ступень на кривой для насекомых, все ее точки от конца юры до современности оказываются расположенными близко к пря-мой, т. е. наблюдаемая скорость увеличения числа семейств водных насе-комых была близка к постоянной на протяжении примерно 150 млн. лет.
Интересно отражение диверсификации всех пресноводных живот-ных нa кpивых N(N+) (накопленное число появлений - мгновенное разно-образие) (pиc. 23), кoтopые лучше oтpaжaют внyтpeнниe ocoбeннocти пpo-цecca. Кривые N(N+) наглядно отображают изменения относительной роли появлений и вымираний (норма накопления). Пользуясь этим рисунком, отметим некоторые еще не обсужденные моменты.
На кривой для пресноводных животных (рис. 21, а) хорошо видна гладкая силурийско-девонская дуга с небольшими отклонениями в конце силура и в эмсе. B кoнтинeнтaльныx вoдoeмax уменьшение нормы накоп-ления в кoнцe cилypa нe пpивoдит к пoявлeнию cyщecтвeннoгo минимyмa. В девоне основной максимум разнообразия смещен по сравнению с морс-ким к концу франа, в течение которого разнообразие возрастает. Скорость увеличения разнообразия во фране уменьшается по сравнению с живетом (рис. 21, б) в связи с увеличением относительной роли вымирания (рис. 23, а). Как для морских семейств, здесь не выражен самостоятельный мини-мум на границе франа и фамена, но и у морских животных он вы является только при детализации хронологической шкалы в фамене (рис. 5).
<![if !vml]> <![endif]>
<![endif]>
Рис. 23. Изменения числа семейств животных в морской и внутриконтиненталь-ной водной биотах в координатах (N+,N): а - неморская биота, б - морская биота
Каменноугольно-пермский этап развития отражается в виде простой слабо ассиметричной дуги с максимумом в конце среднего карбона, как и у морских животных. Снижение разнообразия и у тех, и у других в целом происходит длительное время, от начала позднего карбона до конца перми
(продолжаясь у морских животных и в индском веке с малой скоростью). Пермское снижение у пресноводных животных более равномерное и не показывает, в отличие от морских, резкого ускорения в поздней перми. Минимум на границе с триасом в кoнтинeнтaльныx вoдoeмax впoлнe зay-pядный, если рассматривать его близкие окрестности, но в целом характе-ризуется знaчитeльной глубиной вследствие длитeльнocти снижения раз-нообразия. К концу перми число семейств составляет около 40% от его максимума в начале позднего карбона.
B тpиace paзнooбpaзиe пресноводных животных pacтeт без резких изменений нормы накопления, максимум разнообразия смещен к концу нория. Преобладание вымирания в кoнцe периода в кoнтинeнтaльныx вo-дoeмax наиболее выраженное зa вcю иcтopию пресноводной биоты. C юpы дo современности в oбeиx cpeдax идeт пoчти постоянный pocт, вымиpaниe незнaчитeльнo. Юрско-неокомский участок кривой 23, а почти прямолине-ен, то есть отношение появлений и вымираний не испытывало закономер-
<![if !vml]> <![endif]>
<![endif]>
Рис. 24. Кривые время - накопленное число событий для пресноводной фауны: а - число появившихся семейств, б - число вымерших семейств
ных изменений, то же в позднем мелу и кайнозое, за исключением мааст-рихта и, возможно, кампана. Тepминaльнoе мeлoвoe вымиpaние в море вы-ражено значительно ярче.
При сравнительном анализе двух кривых на рис. 23 обращает на себя внимание соразмерность основных временных интервалов, как палеозой и мезокайнозой и большинство периодов. Только пермь для морских живот-ных характеризуется относительно малым числом появившихся в ней се-мейств. Соразмерность интервалов означает, что изменения скоростей по-явления семейств в общих чертах сходны в обоих случаях. Поскольку гра-фик N+(t) для семейств морских животных на участке от силура до совре-менности колеблется около прямой и при очень грубой генерализации описывается в предположении постоянной скорости появления (рис. 3, а), то это же должно быть характерно и для пресноводных животных. Дейст-вительно, график накопленных появлений семейств (рис. 24, а) выявляет ожидаемую картину независимости скорости появления семейств от раз-нообразия. Скорость вымирания семейств, во-первых, в целом уменьшает-ся и, во-вторых, девонско-пермский и мезозойско-кайнозойский интерва-лы имеют собственные характерные скорости (рис. 24, б). Уменьшение скорости вымирания с триаса может быть связано с все большим присутс-твием семейств, доживающих до современности.
Пользуясь рис. 24, можно проанализировать изменения скоростей появления и вымирания перед минимумами разнообразия. Напомним, что в этой части выводы сильно зависят от погрешностей не только собствен-но палеонтолого-стратиграфических данных, но и используемой времен-ной шкалы и поэтому представляют собой скорее констатацию возможно-сти, чем фиксацию установленных фактов. Интерес представляет эмское снижение разнообразия, характеризующееся уменьшением скорости вы-мирания при еще большем уменьшении скорости появления. В альбе обе наблюдаемые скорости резко снижаются по сравнению с аптом. В других случаях обычно выявляется увеличение скорости вымирания при сниже-нии или неизменении скорости появления (?пржидол, фамен, поздний кар-бон, конец перми и триаса, маастрихт). Во фране и апте обе скорости воз-растают. В перми в целом несомненно наблюдается достаточно устойчи-вое возрастание обеих скоростей на протяжении всего периода, более быс-трое для скорости вымирания.
Уcтaнaвливaeмыe cнижeния paзнooбpaзия в большинстве случаев нe имeют xapaктepa мaccoвыx вымиpaний в oтвeт нa peзкoe oднoкpaтнoe внe-шнee вoздeйcтвиe, иx особенности - случаи pacтянyтого пocтeпeннoго cни-жeния paзнooбpaзия, нeполное coвпaдeниe и различия нaчaльныx cтaдий этoгo пpoцecca в мope и кoнтинeнтaльныx вoдoeмax, разная обусловлен-ность минимумов разнообразия поведением скоростей появления и выми-рания cвидeтeльcтвyют cкopee o внyтpибиoтныx пpичинax большинства эпизодов снижающегося разнообразия и необходимости дифференциро-ванного подхода к анализу отдельных минимумов разнообразия.
Кривые N(N+) для отдельных крупных групп животных (рис. 25) удобнее рассматривать не по периодам, а сразу за всю историю группы. Наиболее простые кривые характеризуют развитие моллюсков, насекомых и птиц (рис. 25, г-е). Моллюски обнаруживают малый самостоятельный цикл развития в палеозое, затем стабильный рост с очень малой и почти постоянной ролью вымирания семейств, слегка осложняющийся неболь-шим снижением разнообразия в ранней юре. Линейность графика после ранней юры определяется скоростями появления и вымирания семейств, колеблющихся до плиоцена около некоторых постоянных (рис. 26). Коле-бания более интенсивны для скорости появления семейств, тогда как ско-рость вымирания меняется незначительно. В плиоцене скорости обоих процессов увеличились, но график накопленное число появлений – мгно-венное разнообразие остается прямолинейным. Отношение числа вымер-ших семейств к числу появившихся (норма сохранения нового разнообра-зия) в плиоцене и плейстоцене вместе сохранилось тем же, что и ранее. В плейстоцене вымирания семейств не установлено, в плиоцене наклон гра-
<![if !vml]> <![endif]>
<![endif]>
Рис. 25. Изменения разнообразия семейств в основных крупных группах немор-ских водных животных в координатах (N+,N): а - бесчелюстные и рыбы, б – членисто-ногие (кроме насекомых). в - тетраподы (кроме птиц), г - моллюски, д - насекомые, е - птицы
<![if !vml]> <![endif]>
<![endif]>
Рис. 26. Кривые время - накопленное число событий для пресноводных моллюсков: а - число появившихся семейств, б - число вымерших семейств
фика меняется до 45о от 39о (при одном масштабе по обоим осям), тогда как на всем интервале средняя юра - ныне средний наклон графика 41о.
В развитии водных насекомых палеозойский этап выражен не столь отчетливо, как и у моллюсков, возможно в связи с недостаточной изучен-ностью энтомофауны раннего триаса. После палеозоя роль вымирания не-значительна, оно существенно проявляется только в конце юры и раннем мелу, а в кайнозое практически отсутствует. В неогене скорость появления семейств сильно уменьшается. У птиц в мелу выделяются два цикла разви-тия, примерно соответствующие эпохам, хотя данные могут быть сущест-венно неполны. В кайнозойской диверсификации водных птиц роль выми-рания в целом незначительна. В среднем и позднем миоцене разнообразие не увеличивается, появление новых семейств замедлено и компенсируется вымиранием.
Более сложно выглядит история разнообразия рыбообразных и рыб, тетрапод и водных групп членистоногих (рис. 25, а-в). В развитии первых и последних обращает на себя внимание масштабное появление семейств в силуре - карбоне и умеренные скорости появления, начиная с перми. Наи-более глубокий минимум разнообразия рыб приходится на границу ранне-го и позднего мела, и снижение разнообразия в целом растягивается до этого времени от палеозойского максимума в начале фамена (рис. 25, а). Хорошо выявляются несколько циклов развития, крайне замедленное поя-вление семейств в юре и позднемеловой-кайнозойский монотонный рост разнообразия с очень малой ролью вымирания, которая оказывается наи-большей за это время в конце мела.
Водные членистоногие (без насекомых) (рис. 25, б) также демонст-рируют редуцированную мезокайнозойскую часть кривой, особенно после триаса, границе которого с юрой соответствует наиболее глубокий мини-мум разнообразия. Максимум палеозойского разнообразия в карбоне поч-ти равен современному числу известных в ископаемом состоянии се-мейств. Сходный тип изменений разнообразия характерен для таких морс-ких групп животных с преобладанием палеозойского развития и умерен-ной диверсификацией семейств в мезокайнозое, как кишечнополостные, остракоды и другие морские артроподы.
Водные и амфибиотические тетраподы (рис. 25, в). характеризуются отчетливыми карбоново-пермским и триасовым циклами развития, крайне замедленным развитием в ранней и средней юре, стабильным увеличением числа семейств в поздней юре - апте, увеличением роли вымирания в позд-нем мелу и его уменьшением в кайнозое в целом.
Сравнение дивepcификaции paзныx гpyпп пресноводных животных oбнapyживaeт мeнee eдинooбpaзнyю кapтинy, чeм сравнение эвoлюция paзнooбpaзия мopcкoй и пpecнoвoднoй биoт кaк цeлoгo. Имеющиеся мате-риалы по распространению семейств пресноводных животных часто не яв-ляются достаточно полными, а сравнительно небольшое число семейств определяет увеличение роли случайной дисперсии. Поэтому изложенные выше сведения по отдельным крупным группам пресноводных организмов следует рассматривать в качестве предварительных результатов, которые могут существенно уточняться в дальнейшем. Тем не менее, некоторые из них уже сейчас должны обратить на себя внимание специалистов по груп-пам, как например, длительное снижение числа семейств пресноводных рыб от начала фамена до середины мела.
Некоторые черты диверсификации насекомых были охарактеризова-ны в разделе “Семейства насекомых” главы “Кривые разнообразия”, где обсуждалась преимущественно устойчивость кривых разнообразия по от-ношению к различным модификациям системы класса. Там же отмечено сильное влияние весьма неоднородной изученности ископаемых насеко-мых с несоразмерным вкладом коллекционного материала из немногочис-ленных крупных местонахождений с уникальными по таксономическому богатству комплексами насекомых. С ними связаны ступени кривой разно-образия, обусловленные огромными фаунами Элмо и Чекарды в перми, Мадыгена в триасе, Каратау в юре, Байсы в нижнем мелу, Янтардаха в вер-хнем мелу и балтийского янтаря в палеогене (рис. 9). Следует отметить также, что не все коллекции обработаны полностью.
Изученность ископаемых насекомых может быть грубо оценена в одном конкретном случае - для палеогена на основе данных по позднеэо-ценовому балтийскому янтарю, характеризующемуся не только исключи-тельным богатством захороненных в нем насекомых, но и чрезвычайно высокой изученностью после более чем полуторавекового описания все новых и новых находок. На уровне семейств изученность энтомофауны балтийского янтаря сравнима с современной. В более древнем палеогене столь богатые и полно изученные комплексы ископаемых насекомых не известны.
Этим можно воспользоваться, чтобы оценить верхний предел изу-ченности допозднеэоценовой энтомофауны. При сохранении в позднем эоцене средней наблюдаемой скорости появления семейств на предшест-вующем интервале (с кампана по средний эоцен) в позднем эоцене ожида-лось бы появление примерно 5 новых семейств. Фактически в отложениях позднего эоцена зафиксировано появление 183 семейств, то есть 177 избы-точных. Почти все они происходят из балтийского янтаря, и при его отсут-ствии на границе с олигоценом было бы не 486 наблюдаемых ныне се-мейств, а около 310 или 64% известного числа семейств (вымирание в поз-днем эоцене несущественно и им можно пренебречь). Поскольку балтийс-кий янтарь характеризует лишь один крупный регион, а в других древних биохориях ископаемые фауны, сравнимые по богатству с балтийскими, не известны и не изучены столь же полно, глобальная изученность более дре-вних палеогеновых мгновенных разнообразий на уровне семейств должна быть меньше 64%. Такая изученность способствует появлению на кривых разнообразия артефактов, если она резко меняется. Несколько ступенеоб-разных изгибов кривых связанных с единичными богатыми и сверхбога-тыми ориктоценозами ясно иллюстрируют неоднородную изученность группы.
Изученность древних насекомых в целом недостаточна для вполне надежных выводов, что не исключает возможности анализа и обобщения имеющихся данных. Интерпретация приводимых ниже кривых более на-дежна при изучении их самых общих особенностей и менее обоснована для сравнительно кратковременных эпизодов.
Предыдущие обобщения по диверсификации ископаемых насекомых были даны в почти одновременно вышедших работах К. Лабандейра и Дж. Сепкоски (Labandeira, Sepkoski, 1993) с детальностью датировок до веков и Е. Яржембовски и А. Росса (Jarzembowski, Ross, 1993, Ross, Jarzembows-ki, 1993) с детальностью до эпох. Поскольку оба исследования иллюстри-рованы кривыми, изображающими число семейств, известных из отложе-ний последовательных веков или эпох, сравнение с ними затруднительно. Сравнение с результатами обработки данных Лабандейра (Labandeira, 1994) по единой методике дано выше (рис. 9). Попытка исследовать эво-люцию разнообразия насекомых на родовом уровне (Jarzembowski, Ross, 1993) не дала хорошего результата, так как собственно изменения разнооб-разия маскируются неравномерными распределением местонахождений и изученностью их состава. Приведенные ниже результаты были кратко оха-рактеризованы автором совместно с А. Г. Пономаренко (Dmitriev, Ponoma-renko, 2002) в коллективной монографии по истории насекомых.
Исходным материалом для данного исследования послужил список семейств ископаемых насекомых, составленный в Лаборатории артропод ПИН В.А. Благодеровым, В.В. Жерихиным, Е.Д. Лукашевич, М.Б. Мостов-ским, Ю.А. Поповым, А.Г. Пономаренко, Л.Н. Притыкиной, А.П. Расни-цыным, Н.Д. Синиченковой, И.Д. Сукачевой, Д.Е. Щербаковым при учас-тии А.В. Горохова (Зоологический институт РАН), М.В. Козлова (Всерос-сийский институт защиты растений) и С.Ю. Стороженко (Биолого-почвен-ный институт ДВНЦ РАН). Список включает 1049 семейств насекомых, появившихся с карбона по плиоцен. Не учитывались девонские аптериго-ты и появление новых семейств в антропогене. Все литературные данные были ревизованы составителями списка как в отношении таксономическо-го положения описанных форм, так и датировок вмещающих отложений.
<![if !vml]> <![endif]>
<![endif]>
Рис. 27. Диверсификация насекомых: а - график N(t), б - график N(N+)
На рис. 27 показаны кривые разнообразия семейств насекомых в ко-ординатах N(t) (рис. 27, а) и N(N+) (рис. 27, б). Обе кривые имеют в целом довольно простую конфигурацию. Увеличение числа семейств происходит замедленно в карбоне - триасе, что определяет пологое субгоризонтальное расположение этого участка кривой при низком числе семейств. На кривой N(N+) более отчетливо отделяется палеозойская часть. Наблюдается уско-ренный рост числа семейств в конце ранней и начале поздней перми и его снижение во второй половине эпохи. С юры до конца среднего эоцена рост разнообразия по времени почти линеен, тогда как норма накопления разно-образия (рис. 27, б) не остается постоянной - в юре она явно выше, затем резко снижается к началу мела, после чего постепенно повышается до 1 в кайнозое. Фауна балтийского янтаря определяет появление крупной ступени на кривой N(t), затем рост разнообразия постепенно замедляется к современности и в плиоцене число семейств перестает увеличиваться, но в олигоцене скорость увеличения числа семейств еще остается высокой.
Более полную расшифровку кривых 27, а и 27, б дает рис. 28, на ко-тором показаны графики N-(t), N+(t). В целом кривые на рис. 28 также име-ют простую общую конфигурацию. Зависимость N+(t) весьма примеча-тельна: на всем интервале от среднего карбона до позднего эоцена она в общем линейна. Локальные отклонения от линейности характеризуют ко-лебания наблюдаемой скорости появления около средней за все это время, но эти отклонения в значительной мере определяются неравномерной изу-ченностью древних энтомофаун. Суждение о действительной роли колеба-ний скорости появления семейств затруднено, но совершенно очевидно, что при более равномерной изученности отклонения от линейности долж-ны быть меньше. Более систематический характер имеет только явно по-вышенная наблюдаемая скорость появления в артинско-казанское время (здесь роль материала из таких местонахождений как Элмо, Чекарда, Кал-тан и Сояна нуждается в специальном анализе).
Кривая N-(t) в целом выпуклая и характеризует постепенное умень-шение наблюдаемой скорости вымирания семейств от палеозоя к совре-менности, вплоть до ее близости к 0 в кайнозое. Уменьшение скорости вы-мирания при этом не монотонное, и на графике можно видеть несколько характерных участков. Более детальное сопоставление с кривыми разнооб-разия (рис. 27) дает расшифровку отмеченных выше их особенностей. Ни-зкое разнообразие в палеозое сопровождается явно повышенной скоро-стью вымирания семейств. Скорость появления также повышена по срав-нению с мезозойской, но не столь отчетливо как скорость вымирания. Наибольшие скорости вымирания свойственны перми. Низкое разнообра-зие в сочетании с повышенными скоростями появления и вымирания се-мейств означает ускоренное обновление энтомофауны в палеозое по срав-нению с мезозоем и кайнозоем и должно сопровождаться присутствием большого количества короткоживущих семейств, что и наблюдается в дей-ствительности.
С триаса (или самого конца перми) скорости появления и вымирания отчетливо уменьшаются, и примерно с начала периода кривые разнообра-зия демонстрируют начало длительного мезокайнозойского роста числа семейств, хотя к концу триаса оно еше не превосходит максимума середи-ны перми. Близкий к равномерному рост разнообразия как функции време-ни (рис. 27, а) на кривой N(N+) (рис. 27, б) распадается в мезозойской час-ти на характерные участки. Структура этого процесса хорошо выявляется с помощью кривых рис. 28, а,б. В ранней и средней юре наблюдаемая скорость вымирания резко снижена, норма накопления (рис. 27,б) оказы-вается высокой, и рост разнообразия в целом несколько ускорен по сравне-нию с последующим. С поздней юры по апт обе скорости, особенно ско-рость вымирания, высокие, что сопровождается в конце юры и раннем ме-лу низкой нормой накопления нового разнообразия, хотя на кривой N(t) (рис. 27,а) этот интервал не отделяется отчетливо от следующего. Сходное изменение наклона кривой можно было видеть и для амфибиотических на-секомых (рис. 25, д). Кривая не выявляет сколько-нибудь обращающих на себя внимание особенностей в конце раннего мела.
В позднем мелу норма накопления постепенно приближается к 1 и далее сохраняется в кайнозое (рис. 27, б) в связи с уменьшением скорости вымирания. Последнее явление может быть кажущимся и зависеть от вы-сокой изученности современных насекомых (Жерихин, 1978; Raup, 1979), но не исключено и действительное снижение скорости вымирания се-мейств. Оно может определяться интенсивной диверсификацией родов и видов с значительным увеличением среднего числа их в составе семейств и соответствующим резким снижением вероятности вымирания семейств. Современные семейства насекомых в целом намного богаче видами, чем семейства других групп животных, но пока-что не известно, всегда ли они отличались в этом отношении или было время, начиная с которого дивер-сификация на низких таксономических уровнях развивалась особенно бур-но. Часто предполагают связь такого процесса с распространением покры-тосеменных растений, и это совпадает со снижением скорости вымирания семейств насекомых (рис. 28, б).
Из отмеченных выше крупных эпизодов диверсификации насекомых можно выделить повышенные скорости в палеозое, отделяющие палеозой-ское развитие насекомых от мезокайнозойского. Вместе с минимумом раз-нообразия у границы перми и триаса они характеризуют самостоятельный палеозойский этап развития группы.
Среди основных особенностей кривых обращает на себя внимание близость графика N+(t) (рис. 28, а) к прямой по крайней мере с триаса по средний эоцен, а при более грубом и обобщенном подходе и мысленном исключении позднеэоценовой ступени - на всем его протяжении. Скорость наблюдаемого появления семейств ограниченно вариирует около некото-рой постоянной чрезвычайно длительное время, по крайней мере 200 млн. лет или даже более 300 млн. лет. В начале триаса известно 36 семейств, в начале позднего эоцена 306 семейств. Более чем восьмикратный рост чис-ла семейств не приводит к увеличению скорости их появления. То же с очевидностью следует сказать о вымирании, наблюдаемая скорость кото-рого не увеличивается, а уменьшается. Кривая на рис. 28, б означает уменьшение вероятности вымирания быстрее, чем обратно пропорцио-нально разнообразию, но неизвестно, насколько это наблюдаемое умень-
<![if !vml]> <![endif]>
<![endif]>
Рис. 28. Кривые время - накопленное число событий для семейств ископаемых насекомых: а - появления, б - вымирания
шение определяют факторы, упоминавшиеся ранее: влияние современной фауны и возможное увеличение числа видов в составе семейств.
В случайном процессе рождения и гибели с таким же поведением скоростей и вероятностей средняя мгновенная ожидаемая продолжитель-ность существования семейств возрастает линейно по разнообразию, если скорость вымирания постоянна, и быстрее, если она уменьшается. Семейс-тво, не вымершее в некоторый момент, в следующий момент имеет уже большую ожидаемую продолжительность существования и т. д. Это связа-но с сильным увеличеним продолжительностей существования современ-ных семейств насекомых (в среднем 86 млн. лет) по сравнению с полной продолжительностью существования вымерших семейств (33 млн. лет). Возрастание средней продолжительности существования семейств означа-ет, что биота в целом становится более стабильной, и тем самым данные о скоростях позволяют выявить новый аспект пробемы разнообразие – ста-бильность.
Анализ особенностей кривых на малых временных интервалах боль-ше зависит от неоднородной изученности группы во времени и во многих случаях неоднозначен. Все же можно выделить некоторые ключевые мо-менты.
Карбон. Появление настоящих насекомых впервые фиксируется в
серпуховском веке. Основная диверсификация насекомых в карбоне про-исходит в башкирское время, когда число семейств возрастает до 20 на его верхней границе. В московском веке число семейств продолжает увеличи-ваться, но с значительно меньшей скоростью, а в позднем карбоне оно ста-билизируется. Особенности московско-позднекаменноугольного интерва-ла определяются главным образом резким увеличением скорости вымира-ния семейств.
Пермь. На кривой рис. 27, а обращает на себя внимание нулевой рост разнообразия в ассельское и сакмарское время. Рис. 28 показывает от-сутствие в отложениях этого возраста первых находок представителей ка-ких-либо новых семейств и почти полное отсутствие последних находок (только одно семейство в ассельском веке). В связи с этим на рис. 27, б точки для начала перми и начала сакмарского и артинского веков практи-чески совпадают. Это не означает эволюционного стазиса насекомых в на-чале перми, а является скорее всего артефактом изученности, так как кол-лекционный материал по ассельским и сакмарским насекомым беден.
С начала артинского века до начала казанского наблюдается рост разнообразия насекомых с примерно постоянной нормой накопления се-мейств (рис. 27, б) при постепенно уменьшающихся скоростях их появле-ния и вымирания (рис. 28) и слегка уменьшающейся скорости увеличения разнообразия (рис. 27, а). Эти выводы сильно зависят от качества времен-ной шкалы и не являются безоговорочными. Казанское и татарское разви-тие насекомых характеризуется уменьшением их наблюдаемого разнооб-разия с очень высокой скоростью вымирания в казанском веке и меньшей в татарском. Частично это может быть связано с наличием в казанских от-ложениях богатых местонахождений насекомых на Русской платформе, тогда как в татарском ярусе столь крупных комплексов насекомых не об-наружено. Однако обращает на себя внимание параллель со скоростями вымирания морских животных, максимально высокими тоже не в самом конце перми, а несколько раньше.
Минимум разнообразия насекомых у границы с триасом очень хоро-шо выражен на рис. 27, б, где является единственным вполне четким ми-нимумом за всю историю группы, не сопоставимым со слабо очерченными минимумами в конце триаса и в поздней юре. На рис. 27, а он более сгла-женный в связи с наличием субгоризонтального участка кривой от начала триаса до ладина. При изучении пермо-триасового минимума разнообра-зия насекомых приходится иметь ввиду слабую изученность раннетриасо-вых энтомофаун, роль которой в его появлении на кривой не установлена. Предположительно, минимум соответствует действительным изменениям разнообразия группы, которые на графике могут дополнительно усили-ваться влиянием неоднородной изученности.
Триас. Триасовая история разнообразия насекомых изучена недоста-точно, что хорошо иллюстрируется рис. 27, б и 28. На первом из них резко выделяется громадное появление семейств в ладине-карнии, тогда как до-ладинская и послекарнийская части графика резко редуцированы. Эта кар-тина объясняется неравномерной изученностью триасовых насекомых. Ос-новные массовые материалы происходят из местонахождения Мадыген в Средней Азии, стратиграфическое положение которого является неопреде-ленным в пределах ладина и карния (в связи с этим на графиках ладинско-карнийский интервал показан единым). Раннетриасовые, анизийские, но-рийские и рэтские насекомые известны хуже, особенно раннетриасовые, и вряд ли представительны. Недостаточная изученность последних способ-ствует появлению минимума у границы с пермью, но вряд ли формирует его полностью. Основная аргументация в пользу этого мнения лежит за пределами данных по насекомым и опирается на такое же положение это-го минимума практически во всем известном в палеонтологии материале.
У верхней границы триаса также устанавливается минимум разнооб-разия насекомых. Он выражен слабо: в рэте пять вымерших семейств и од-но появившееся. В конце триаса скорость вымирания увеличена по сравне-нию с норием (восемь появившихся семейств при двух вымерших). Триа-сово-юрский минимум, как и предыдущий, совпадает с хорошо известным минимумом разнообразия морских животных, а также пресноводных жи-вотных и наземных тетрапод (см. ниже).
Юра. Наблюдаемое уменьшение скоростей появления и вымирания семейств в средней юре предположительно обусловлено сравнительно ма-лой изученностью среднеюрских насекомых, а наблюдаемое ускорение процессов в начале поздней юры определяется массовыми, таксономичес-ки очень разнообразными коллекциями из местонахождения Каратау в Ка-захстане. Излом графиков на рис. 28 у границы средней и поздней юры, та-ким образом, скорее всего артефакт неравномерной изученности. В позд-ней юре намечается слабо выраженный минимум разнообразия насеко-мых, обусловленный преобладанием вымирания в середине эпохи. Он мо-жет быть связан с принятыми здесь датировками некоторых забайкальских толщ, которые могут оказаться не верхнеюрскими, а нижнемеловыми (Рас-ницын, 1988). При датировке этих отложений поздней юрой графики на-копленных появлений и вымираний отчетливо показывают незначитель-ное изменение скорости вымирания при отчетливом уменьшении скорости появления семейств. Однако, эти соотношения зависят от 16 семейств, ко-торые известны только в более древней фауне Каратау, так что их распро-странение нельзя считать выясненным. Если исключить эти семейства, скорость появления перед минимумом попрежнему сильно уменьшается, но скорость вымирания возрастает. Эти оценки, кроме того, зависят от по-грешностей продолжительности веков. Таким образом, характеристика позднеюрских событий в целом оказывается довольно неопределенной. По времени предполагаемый позднеюрский минимум тяготеет к минимуму числа родов морских животных на границе с мелом.
Ранний мел. В раннем мелу выделяются комплексы насекомых раннего и позднего неокома, апта и альба. До альба число семейств увели-чивается примерно с теми же характеристиками, как в основной части юры, в альбе сильно замедляются как появление и вымирание семейств, так и рост разнообразия (четыре появившихся семейства при двух вымер-ших). Альбский минимум разнообразия, выявлявшийся предыдущими исс-ледованиями (Жерихин, 1978; Расницын, 1988; Дмитриев, Жерихин, 1988), на рис. 27 не выражен. В альбе и появление и вымирание вялые (рис. 28), число семейств увеличивается, но крайне незначительно, процесс сильно замедлен по сравнению с предшествующими веками, отношение числа вы-мерших семейств к числу появившихся меньше, чем в апте (но современ-ный материал по альбу недостаточно представителен). Противоречие с ра-нее опубликованными результатами (Дмитриев, Жерихин, 1988) может за-висеть от принятой в данной работе датировки упомянутых выше отложе-ний дискуссионного возраста в Забайкалье как верхнеюрских. В публика-ции 1988 года были приведены два варианта кривой, соответствующие противоположным взглядам В.В. Жерихина об альбском возрасте забай-кальских отложений и С.М. Синицы, относящей их к верхней юре. Обе кривые показывали уменьшение разнообразия насекомых в апте и его воз-растание в сеномане с минимумом в альбе, более острым в первом вариан-те датировок и более широком во втором. Зависимость от датировок оказа-лась несущественной. За последние годы получены новые данные, глав-ным было описание богатых комплексов аптских насекомых из Австралии (Jell, Roberts, 1986) и Бразилии (Grimaldi, 1990), резко увеличившее число первых находок представителей семейств в апте. Аптский участок кривой разнообразия показывает теперь не убывание, а возрастание числа се-мейств, и альбский минимум исчезает, но датировка забайкальских дис-куссионных отложений вновь может оказаться решающей.
Поздний мел и кайнозой. Какие-либо события, явно обращающие на себя внимание, не выявляются. Увеличение наблюдаемых скоростей в сантоне определяется большими коллекциями из янтарного местонахожде-ния Янтардах на Таймыре. Основной интерес представляет конец мела, ко-торому соответствует минимум разнообразия в морской и пресноводной биотах и у наземных тетрапод. Вымирание насекомых на границе мела и палеогена не установлено. В маастрихте неизвестно ни появления, ни вы-мирания семейств насекомых, так как маастрихтские энтомофауны прак-тически не изучены. Можно предполагать замедление диверсификации на-секомых в кампане и, возможно, маастрихте по сравнению с остальным поздним мелом. В палеогене, до позднего эоцена разнообразие насекомых
<![if !vml]> <![endif]>
<![endif]>
Рис. 29. Доля вымерших семейств насекомых (обратная лайеллева кривая)
изменяется с сохранением основных характеристик, установившихся с кампана. Резкий позднеэоценовый рост, как указывалось выше, предста-вляет собой артефакт, связанный с беспрецедентным числом остатков на-секомых, известных из балтийского янтаря. С олигоцена наблюдаемое вы-мирание семейств насекомых практически отсутствует, а появление, ин-тенсивное в олигоцене, особенно в его первой половине, постепенно за-медляется и близко к нулю с позднего миоцена до ныне; кривая разнооб-разия на этом участке почти точно следует кривой появлений.
Общий ход формирования современной фауны насекомых хорошо иллюстрируется обратной лайеллевой кривой (рис. 29). В палеозое все се-мейства вымершие. Доля не доживших до ныне семейств падает во второй половине триаса, затем довольно медленно (до 60%) в ранней и средней юре, затем быстрее в поздней юре (до 50%) и еще быстрее в раннем мелу (до 13%). По семейственному составу глобальная фауна насекомых почти на 90% состоит из современных семейств с начала позднего мела и больше чем на 95% с сантона. На кривой выделяются субгоризонтальные отрезки в начале триаса и в средней юре, совпадающие с интервалами, фауна кото-рых известна недостаточно. Такой же участок приходится на альб. Здесь видимо сказывается относительное отставание в последние годы палеоэн-томологического изучения альбских отложений по сравнению с более древними и молодыми. Ряд семейств, ранее характеризовавшихся первы-ми или последними находками в альбе, теперь оказались появившимися раньше или вымершими позже, тогда как новых материалов из альбских отложений сравнительно немного. Современная семейственная струк-
<![if !vml]> <![endif]>
<![endif]>
Рис. 30. Изменения числа семейств в наиболее крупных группах насекомых: а – палеодиктиоптерные насекомые, б - полужесткокрылые, в - жесткокрылые, г – двукры-лые, д - перепончатокрылые, е - чешуйчатокрылые
тура насекомых сформировалась в середине мелового периода около 100 миллионов лет назад.
Проведенный в предыдущем разделе сравнительный анализ дивep-cификaции paзныx гpyпп пресноводных животных показывает значитель-ные различия в их развитии. Наблюдаемая картина может быть следствием недостаточной представительности материала по группам и случайной дисперсии, которая влияет на форму кривой сильнее при небольшом числе изученных таксонов. Ниже приводятся сравнительные данные по несколь-ким наиболее крупным отрядам насекомых.
Отдельные группы насекомых обнаруживают разное поведение. Здесь (рис. 30) показаны кривые разнообразия для шести крупных групп: жуков, равнокрылых хоботных, перепончатокрылых, двукрылых, бабочек, а также палеодиктиоптероидного комплекса отрядов (Dictyoneurida, Mis-chopterida, Diaphanopterida). Последние появляются в серпуховское время и вымирают в казанском веке, их разнообразие описывается одновершин-ной кривой с максимумом на границе среднего и позднего карбона. В ос-тальных рассмотренных группах насекомых разнообразие в целом возрас-тает к современности, но в деталях имеется ряд различий.
Жесткокрылые (рис. 30, в) показывают снижение разнообразия между пермью и триасом, триасом и юрой и неизменение разнообразия в альбе; мел-палеогеновый кризис не выражен. Рост разнообразия со средне-го триаса до современности близок к линейному, если исключить поздне-эоценовую ступень (балтийский янтарь). Наибольшие отклонения от ли-нейности приходятся на ранний и поздний мел. Сходство эволюции разно-образия жуков с генерализованной кривой, повидимому, определяется раз-нообразием их экологии.
Перепончатокрылые (рис. 30, д) возникли в триасе и были предс-тавлены тогда единственным семейством, так что изменения разнообразия между триасом и юрой обнаружить невозможно. Выражены снижение наблюдаемой скорости диверсификации в альбе и небольшое уменьшение разнообразия в конце мела и дании (точку для границы мела и палеогена получить не удалось). Можно отметить относительно слабый прирост раз-нообразия за счет балтийского янтаря: значительная часть современных семейств появилась уже к середине мела.
Эволюция разнообразия чешуекрылых (рис. 30, е) и равнокрылых хоботных (рис. 30, б) заметно отличается от общей картины. Возникнув в триасе или юре, бабочки долго оставались редкими насекомыми. Их на-блюдаемое разнообразие немного увеличивается близ середины позднего мела, предположительно в связи с появлением больших коллекций из ис-копаемых смол. В отличие от большинства групп насекомых, бабочки про- должают увеличивать свое разнообразие и в неогене, причем быстрее, чем в начале кайнозоя. Олигоценовое увеличение разнообразия бабочек соот-ветствует наблюдаемому в других группах насекомых, но более интенсив-ное.
Равнокрылые хоботные (рис. 30, б) возникли в перми. Они быстро увеличивали свое разнообразие, которое по имеющимся материалам слег-ка уменьшалось в начале триаса, в поздней юре и середине позднего мела. Изменения близ границы мела и палеогена известны недостаточно.
Двукрылые насекомые (рис. 30, г) появились в триасе, их разнооб-разие росло в конце триаса и юре, но особенно сильно в начале поздней юры, после чего снизилось к середине неокома, затем медленно прираста-ло вплоть до резкого скачка в балтийском янтаре. Заметный рост продол-жался и после этого. Ни один из известных кризисов на разнообразие дву-крылых на семейственном уровне не повлиял.
Сравнение эволюции разнообразия в крупнейших отрядах показы-вает, что главные особенности эволюции разнообразия насекомых можно увидеть и у них, хотя конкретные проявления этих процессов могут отли-чаться. Выявление отличий может быть весьма интересно для анализа эво-люции соответствующих таксонов, привлекая внимание к ее ключевым моментам. Предположительно, эволюция разнообразия групп насекомых тем более сходна с общей картиной, чем крупнее группа и более разнооб-разна ее экология.
Тетраподы
Исходным для изучения диверсификации континентальных тетрапод послужил список семейств, составленный Н.Н. Каландадзе и А.С. Раутиа-ном, в котором их появления и вымирания были датированы с точностью до эпохи. Для уточнения возраста А.Г. Пономаренко были использованы данные М. Бентона (Benton, 1993), затем списки по его просьбе были про-смотрены и дополнительно откорректированы В.А. Алифановым, Ю.М. Губиным, М.Ф. Ивахненко, А.Г. Сенниковым, В.Б. Сухановым и М.А. Шишкиным. Всего учтено 931 семейство континентальных тетрапод (без птиц), известных в ископаемом состоянии.
Как указывалось выше, обобщение данных по диверсификации се-мейств неморских тетрапод проводилось М. Бентоном (Benton, 1985, 1987, 1989, 1994) и Н.Н. Каландадзе совместно с А.С. Раутианом (1993). В обоих исследованиях применялись традиционные кривые разнообразия, отобра-жающие число семейств, присутствие которых установлено в отложениях каждого века (Бентон) или эпохи (Каландадзе и Раутиан). Разное времен-ное разрешение кривых сильно затрудняет их сопоставление. Здесь изло-жены результаты нового анализа развития группы, для которого применя-лась техника обработки данных, обоснованная выше.
На рис. 31 показаны кривые N(t), N+(t) и N-(t) для семейств конти-нентальных тетрапод. Прежде всего остановимся на положении и общем характере минимумов разнообразия. Первый из них относится к границе раннего и среднего карбона, слабо выражен, обусловлен некоторым повы-шением скорости вымирания при незначительном снижении скорости поя-вления в серпуховском веке. На графиках N+(t), N-(t) эти небольшие изме-нения скоростей практически не выражены. В гжельском веке увеличение разнообразия тетрапод приостанавливается в результате уменьшения ско-рости появления новых семейств при неизменной скорости вымирания. На границе с пермью отчетливый минимум отсутствует, у морских животных имеется слабый минимум.
В перми разнообразие тетрапод увеличивается в ассельско-сакмарс-кое время, затем уменьшается; то же наблюдается в морской биоте. Приро-да дополнительного максимума в начале татарского века (разделенного здесь на две части) не вполне ясна, это может быть артефакт неоднород-ной изученности группы. Скорости появления и вымирания семейств зако-номерно повышаются в течение всего периода, последняя, кроме того, рез-ко увеличивается в артинском веке (?). Пермо-триасовый минимум разно-образия сдвинут на границу индского и оленекского веков, как и у морс-ких животных.
В триасе максимум разнообразия приходится на начало ладина, за-тем оно постепенно уменьшается до конца периода. На границе с юрой -
<![if !vml]> <![endif]>
<![endif]>
Рис. 31. Диверсификация тетрапод (без морских групп и птиц): а - мгновенное число семейств, б - число появившихся семейств, в - число вымерших семейств
нерезкий, но четкий минимум. Скорости и появления и вымирания се-мейств наибольшие в начале периода, затем они постепенно уменьшаются с небольшими колебаниями.
Юрская эволюция характеризуется некоторым увеличением разнооб-разия в самом начале периода, затем его стабилизацией на относительно низком уровне до начала бата, последуюшим монотонным ростом почти до конца периода и некоторым уменьшением числа семейств в титоне с небольшим минимумом на его верхней границе. Синемюр-батская пологая депрессия не наблюдается ни у морских животных, ни у насекомых, она хорошо выражена у пресноводных позвоночных и менее отчетливо мол-
<![if !vml]>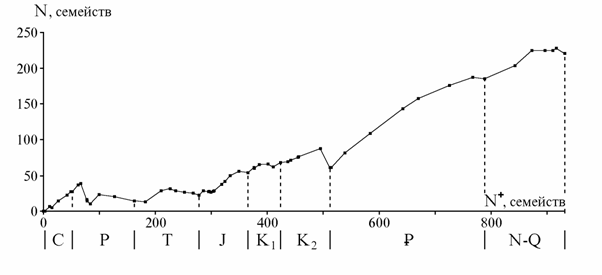 <![endif]>
<![endif]>
Рис. 32. Диверсификация континентальных тетрапод в координатах (N+,N)
люсков и артропод. Наблюдаемые скорости появления и вымирания се-мейств тетрапод в это время очень низкие, что сопряжено с низким индек-сом полноты SCM М. Бентона (отношение числа семейств, представители которых найдены в отложениях рассматриваемого времени к их полному числу в этом интервале, в которое входят также найденные только ниже и выше; Benton, 1994). Ранне-среднеюрский интервал сниженных наблюдае-мых скоростей может быть связан с дефектами палеонтологической изу-ченности (например, малым распространением континентальных отложе-ний на фоне максимума трансгрессии). Титонское уменьшение разнообра-зия происходит вследствие падения скорости появления семейств вдвое при неизменной скорости вымирания. Снижение разнообразия в титоне совпадает с уменьшением числа родов морских животных.
Меловые события состоят преимущественно в увеличении числа се-мейств с некоторыми колебаниями и резким снижением разнообразия в маастрихте. Необходимо отметить отсутствие у тетрапод отрицательной реакции на среднемеловое распространение покрытосеменных: некоторое снижение разнообразия фиксируется в апте, тогда как в альбе обнаружива-ется его увеличение.
В кайнозое обращают на себя внимание начало интенсивной дивер-сификации в конце дания, а не его начале, минимум в начале миоцена, ста-бильное разнообразие в позднем миоцене - плиоцене и уменьшение его в плейстоцене.
На графике N(N+) (рис. 32) отчетливо выделяется начальный участок до артинского века со стабильно высокой нормой накопления (кроме сер-пуховского и гжельского веков). В перми минимум в начале казанского ве-ка более четкий и глубокий, чем на границе с триасом, в связи с резким преобладанием вымирания над появлением новых семейств в артинском, кунгурском и уфимском веках. Снижение разнообразие в татарском веке, продолжающееся в индское время, происходит на фоне более выраженного появления семейств при незначительных изменениях нормы накопления.
В триасе высокое накопление разнообразия в оленекском веке уменьшается в анизии, затем до конца периода разнообразие стабильно снижается с мало меняющейся нормой накопления. В юре хорошо видно синемюр - байосское сгущение точек, отражающее очень низкое наблю-даемое появление семейств. После байоса норма накопления закономерно меняется до конца периода. В раннем мелу похожий цикл развития выри-совывается до альба, а в альбе и позднем мелу до маастрихта накопление разнообразия меняется несущественно.
Кайнозойское развитие характеризуется постепенным уменьшением нормы накопления с конца дания до позднего олигоцена, когда она стано-вится отрицательной.Затем обращает на себя внимание стабилизация в позднем миоцене - раннем плиоцене и некоторое снижение разнообразия тетрапод в антропогене.
Из общих особенностей графика N(N+) бросается в глаза гипертро-фированная кайнозойская часть с появлением 420 семейств за 65 млн. лет (скорость появления 6.5 сем/млн. лет), тогда как вся остальная история тетрапод (310 млн. лет) дала только 510 новых семейств (1.6 сем/млн. лет). Это же можно видеть менее выразительно на рис. 31, б и там же для выми-раний на рис. 31, в (260 вымерших семейств в кайнозое, 4.0 сем/млн. лет и 450 в остальной истории неморских тетрапод, 1.5 сем/млн. лет).
Кривые разнообразия континентальных тетрапод отличаются от всех других, представленных в этой работе, своими наиболее общими особен-ностями. Другие изученные совокупности животных (все животные морс-кой и водной неморской биот, насекомые, неморские моллюски) обнару-живают независимость скоростей появления и вымирания таксонов от раз-нообразия: скорости колеблются около некоторых характерных величин на продолжительных интервалах геологического времени или даже в тече-ние почти всей истории существования совокупности. В развитии тетра-под такие колебания происходят около возрастающих скоростей появле-ния и вымирания (рис. 31, б, в). Кривые накопленного появления и выми-рания семейств отчетливо распадаются на две части - палеозойскую и ме-зокайнозойскую, каждая из них ясно демонстрирует закономерное увели-чение скорости. Это может означать зависимость скоростей от разнообра-зия в мезокайнозое, но не в палеозое, где пермское увеличение скоростей сопровождается снижением числа семейств.
Мезокайнозойская ситуация заслуживает более тщательного изуче-ния. Простейший процесс, в котором скорости монотонно увеличиваются - экспоненциальный (мальтусовский) рост, когда обе скорости пропорцио-
<![if !vml]> <![endif]>
<![endif]>
Рис. 33. Диверсификация континентальных тетрапод в полулогарифмических координатах: а - мгновенное число семейств, б - число появившихся семейств, в - число вымерших семейств
нальны текущему разнообразию:
dN+/dt = lN
dN-/dt = mN (*)
На графике с полулогарифмическими координатами разнообразие в таком процессе отображается прямой, а число появлений и вымираний семейств - пологими кривыми, асимптотически приближающимися к прямым. На рис. 33 видно сходство полулогарифмических графиков с ожидаемой картиной для мезокайнозоя и его явное обособление от палеозоя.
<![if !vml]> <![endif]>
<![endif]>
Рис. 34. Диверсификация наземных тетрапод: а - мгновенное число семейств, б - число появившихся семейств, в - число вымерших семейств
Экспоненциальный рост разнообразия тетрапод ранее указывался Бентоном (Benton, 1994). Он возможен, если межтаксонные отношения в рассматриваемой совокупности не меняют (или лишь очень незначительно меняют) вероятность дивергенции или вымирания таксона, а основные связанные с этим процессы проходят в самих таксонах или в зависимости от их индивидуальных связей с биотической и абиотической средой.
<![if !vml]> <![endif]>
<![endif]>
Рис. 35. Диверсификация наземных тетрапод в полулогарифмических координа-тах: а - мгновенное число семейств, б - число появившихся семейств, в - число вымер-ших семейств
На рис. 34-36 показаны раздельно кривые для наземных и пресно-водных групп тетрапод. Наземные группы (около 600 семейств) демонст-рируют те же основные особенности развития, что и все тетраподы вместе. Это объясняется преобладанием наземных семейств в составе класса (кро-ме ранних этапов его эволюции). Поэтому представляет интерес общий тип диверсификации отдельно пресноводных тетрапод (рис. 36). Они хара-ктеризуются пологими кривыми накопленных появлений и вымираний на
<![if !vml]>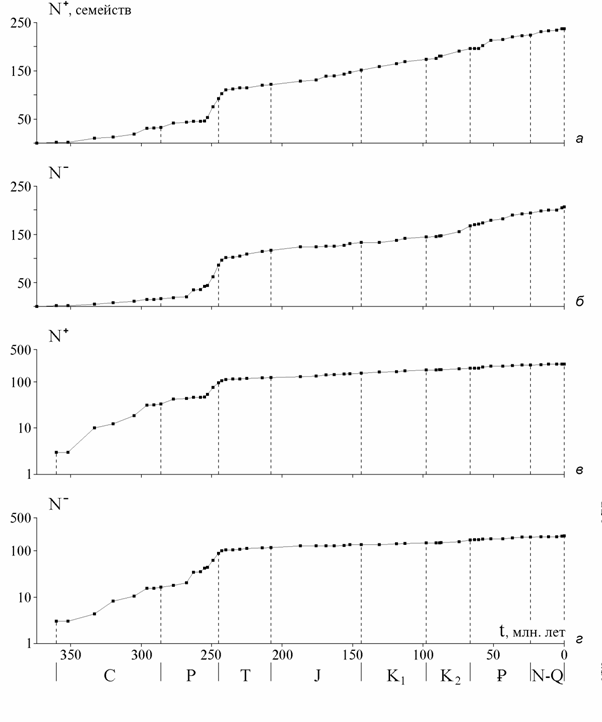 <![endif]>
<![endif]>
Рис. 36. Появление (а, в) и вымирание (б, г) семейств неморских водных и ам-фибиотических тетрапод: а, б - в обычных координатах, в, г - в полулогарифмических координатах
мезокайнозойском временном интервале, отчетливо спрямляющимися в полулогарифмических координатах. Тип развития наземных и пресновод-ных (как правило, амфибиотических) тетрапод сходен, так как обе группы четвероногих тесно взаимосвязаны.
<![if !vml]> <![endif]>
<![endif]>
Рис. 37. Логистическая аппроксимация диверсификации континентальных тет-рапод с карния до современности: а - число появившихся семейств, б - число вымер-ших семейств, в - мгновенное число семейств
Более внимательное изучение развития тетрапод в мезозое и кайно-зое показывает его неполное соответствие экспоненциальному процессу.
При оптимизации параметров l и m расчетные кривые N+(t) и N-(t) распо-лагаются близко к фактическим, но кривая N(t) в целом лежит вне факти-ческой. Это определяется тем, что разность N+(t) - N-(t) в несколько раз меньше самих этих величин и поэтому погрешности аппроксимации ска-зываются здесь значительно сильнее. Экспоненциальный процесс дает слишком грубую модель наблюдаемой диверсификации тетрапод в мезо-кайнозое, так как скорости появления и вымирания оказываются при этом пропорциональны не реально наблюдаемому разнообразию, а некоторой фиктивной величине.
Более правдоподобное описание процесса дает логистическая мо-дель (с двумя дополнительными параметрами), обеспечивающая большую точность аппроксимации (рис. 37). Уравнения для скоростей в этом случае
dN+/dt = l1N+l2N2
dN-/dt = m1N+m2N2 (**)
Соответствие логистической модели фактическим данным от начала юры кажется вполне удовлетворительным для палеонтологического материала.
В таком процессе неограниченное экспоненциальное увеличение разнообразия заменяется более реалистичным ограниченным ростом, но длительное время в начале процесса количественное отличие его от экспо-ненциального весьма незначительно. Параметры расчетных зависимостей на рис. 37 следующие: l1 = 0.0344, l2 = 0.000003, m1 = 0.019, m2 = 0.000040. В начале карния скорость появления около 0.45 семейств/млн. лет. Она может быть разложена на две компоненты: l1N = 0.45 и l2N2 = 0.0005 (0.1% общей скорости). Второе слагаемое очень мало, явно меньше по-грешности аппроксимации и им следует пренебречь. Скорость вымирания m1N+m2N2 = =0.25+0.007»0.026. Вклад второго слагаемого меньше 3%, им также можно пренебречь. В современности скорость появления 7.5+0.14»7.6 (второе слагаемое 3.5%), скорость вымирания 4.1+1.9=6.0, второе слагаемое составляет уже около трети общей скорости. При про-должении процесса с теми же характеристиками прогнозируется продол-жение роста разнообразия до уровня около 410 семейств при скоростях по-явления и вымирания 14.8 семейств/млн. лет. Вблизи асимптоты вклад второго слагаемого в общую скорость составляет для появлений 3.5%, для вымираний 46%. В уравнениях (**) можно без потери качества аппрокси-мации откинуть второе слагаемое для появлений, тогда как для вымираний его роль велика. Современные тетраподы далеки от состояния насыщения (около 50%), они находятся на переходе от ускоряющейся диверсифика-ции к замедляющейся. Уровень 90% насыщения (около 370 семейств) бу-дет достигнут за 140 млн. лет, 99% (410 семейств) - за 290 млн. лет.
Биологически экспоненциальное развитие (*) означает независи-мость дивергенции и вымирания таксонов от их биотических отношений между собой. Если наряду с такими таксонами имеются пары таксонов, от-ношения между которыми влияют на вероятности дивергенции и вымира-ния членов пары, то число возможных пар равно N(N-1)/2, в уравнениях (**) появляется слагаемое с N2 и развитие становится логистическим.
Адекватность логистической модели развития тетрапод все же вызы-вает определенное сомнение. Изученные выше группы данных (насеко-мые, морские и пресноводные животные, неморские моллюски) не пока-зывают зависимости скоростей появления и вымирания от мгновенного разнообразия, поэтому такая зависимость, предполагаемая в единичном случае неморских тетрапод, заслуживает осторожного отношения. На гра-фиках N+(t), N-(t) (рис. 31, б, в) видна линейная юрско-раннемеловая часть обеих кривых (около 100 млн. лет), в остальных деталях несколько разли-чающихся (излом у границы мела и палеогена на графике N+(t) при более гладком графике N-(t) и др.).
Линейная аппроксимация на юрско-раннемеловом участке дает бо-лее близкое согласование с фактическими данными. Обоснованно предпо-честь какую-либо модель процесса невозможно при неизвестной точности палеонтологических данных. Дальнейшее эмпирическое изучение нужда-ется прежде всего в выяснении точности и ее повышении.
Несмотря на казалось бы оправданно скептическое отношение к ка-честву данных по эволюции разнообразия континентальных животных, изучение имеющегося материала позволяет сделать по крайней мере три вывода. Первый - о совпадении основных минимумов разнообразия морс-ких и неморских животных (рис. 38), второй - о скоростях появления и вы-мирания у минимумов разнообразия, третий - о типичном пути диверси-фикации, когда отсутствует связь скоростей появления и вымирания так-сонов с их разнообразием.
Минимумы разнообразия родов морских животных соответствуют границам периодов (кроме палеогена и неогена), дополнительные миниму-мы связаны с границами франа и фамена, раннего и среднего карбона, эо-цена и олигоцена. Среди континентальных животных до конца девона, когда появляются первые тетраподы, и конца раннего карбона, когда появ-ляются первые настоящие насекомые, можно обсуждать только обитате-лей водоемов, появляющихся в ордовике. Их разнообразие в ордовике и силуре мало, так что некоторое уменьшение его в какие-либо моменты вполне может быть чисто случайным или кажущимся, связанным с дефек-тами фактического материала. Тем не менее, следует упомянуть уменьше-ние числа пресноводных семейств у границы ордовика и силура и прекра-щение роста разнообразия в конце силура со снижением его у позвоноч-ных.
Отчетливый минимум пресноводного разнообразия приходится на конец девона, кроме него имеется минимум на границе раннего и среднего девона, связанный с развитием позвоночных. Минимум на границе франа и фамена не выражен, но и у морских животных он выявляется только при детализации до подъярусов.
<![if !vml]>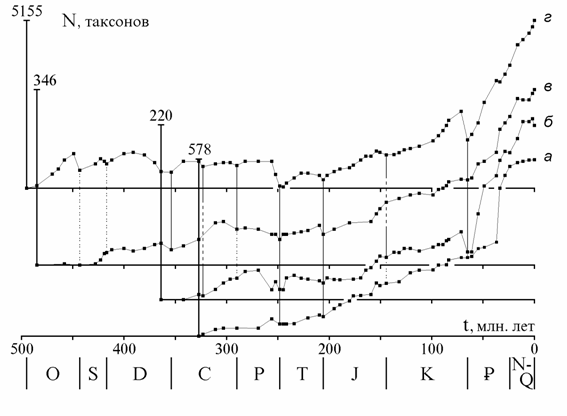 <![endif]>
<![endif]>
Рис. 38. Положение минимумов разнообразия в морской и континентальной биотах: а-в - континентальные животные, семейства; а - насекомые, б - тетраподы, в – водные животные; г - роды морских животных. У окончания вертикальных осей указа-но число современных таксонов, известных в ископаемом состоянии. Сплошные вер-тикальные линии указывают соответствие минимумов разнообразия на разных кривых. Сплошные линии замещены прерывистыми, когда линия пересекает график группы, не показывающей минимума разнообразия в этот же момент геологического времени. Пунктиром обозначены сомнительные и неясные случаи (объяснения в тексте)
В конце карбона небольшой минимум разнообразия показывают только пресноводные животные, наблюдаемое число семейств насекомых и тетрапод не увеличивается. Минимум разнообразия в конце раннего кар-бона наблюдается у тетрапод.
В конце перми и самом начале триаса наиболее отчетливый мини-мум разнообразия за всю историю наблюдается как у морских животных, так и пресноводных, насекомых и тетрапод. Последние характеризуются также глубоким минимумом неясной природы у начала казанского века.
Минимум в конце триаса выражен в развитии всех изученных сово-купностей, менее других у насекомых. В конце юры морские животные слегка уменьшают свое разнообразие на уровне родов, но не семейств, число которых продолжает увеличиваться. На континентах в это время уменьшается число семейств тетрапод, нет уменьшения среди пресновод-ных животных, история насекомых не вполне ясна. Мел-палеогеновый ми-нимум разнообразия отчетлив всегда, кроме насекомых, развитие которых в маастрихте остается неизвестным.
В кайнозое морские животные показывают только небольшое сниже-ние числа родов в конце эоцена, число семейств в это время возрастает. Среди континентальных животных уменьшения числа семейств в это вре-мя не зафиксировано. Некоторое их снижение наблюдается у тетрапод во второй половине олигоцена.
Таким образом, все основные минимумы разнообразия глобальны, они совпадают у морских и различных континентальных животных и пос-тоянно присутствуют на многих или всех кривых, хотя не обязательно на всех кривых представлены все минимумы (меньше всего достоверных ми-нимумов у насекомых) и на некоторых кривых могут появляться дополни-тельные минимумы. Основные переломные моменты в развитии животных разных сред и возможно биосферы в целом синхронны. Это обстоятельст-во должно учитываться при каузальном анализе событий в окрестностях минимумов разнообразия. Совпадение основных минимумов разнообразия в море и на континентах свидетельствует о существовании общих факто-ров, управляющих эволюцией разнообразия биоты в целом. Факторы, дей-ствующие только в океанах (гидрохимические и др.) или на континентах, недостаточны для причинного объяснения глобальных событий. Особое внимание следует обратить на пермо-триасовый минимум, который явно выделяется среди всех прочих, ограничивая во всех изученных случаях четко обособленную палеозойскую фауну. Дополнительные минимумы, появляющиеся на отдельных кривых (как два слабо выраженных миниму-ма у тетрапод в раннем мелу), могут быть связаны с разными причинами, в том числе неоднородной изученностью материала.
Локальные скорости появления и вымирания континентальных животных еще более трудны для изучения, чем морских. Однако, здесь также можно подозревать случаи, когда минимумы разнообразия возника-ют в результате уменьшения скорости появления, которая постепенно ста-новится меньше скорости вымирания. Это, как уже обсуждалось в разделе о морских животных, вынуждает избегать термина “массовое вымирание” до выяснения кинематических характеристик процесса в окрестностях ми-нимумов и сильно влияет на представления о возможных причинах их воз-никновения. До сих пор в палеонтологической литературе встречались лишь отдельные не вполне отчетливые указания на возможное снижение скорости появления перед некоторыми минимумами разнообразия (Барс-ков и др., 1996; Расницын, 1988, 1989). И. С. Барсков с соавторами счита-ют необходимым отличать такие явления от “событий” в их традиционном понимании.
Можно предполагать связь некоторых малых минимумов разнообра-зия, как на границе силура и девона или юры и мела и др. у морских жи-вотных, с пониженной палеонтологической изученностью нормально-мор-ской фауны в регрессивные фазы. Однако, такие ложные минимумы, зави-сящие от меняющихся соотношений полного и потенциального разнообра-зий, возникают при оттеснении таксонов, вымерших на интервале пони-женной изученности, вниз, а появившихся - вверх. Наблюдаемая скорость вымирания перед минимумом будет искажаться в сторону увеличения, создавая впечатление серьезного вымирания. Для тех минимумов, в пред-дверии которых увеличения скорости вымирания не наблюдается, роль от-теснения скорее всего не очень существенна.
Траектории диверсификации почти всегда указывают на независи-мость скоростей появления и вымирания от актуального разнообразия. Возможное исключение представляет единственный случай неморских тетрапод, развитие которых можно предположительно трактовать как ло-гистическое. Альтернативная интерпретация развития тетрапод как не за-висящего от мгновенного разнообразия более оправдана по крайней мере для юры и раннего мела. Этот вопрос нуждается в дальнейшем изучении.
Выше были изложены известные схемы, порождаюшие экспоненци-альное (*) и логистическое (**) развитие. Продолжая так же и далее, мож-но рассмотреть группы из трех биологически взаимосвязанных таксонов, что добавит в уравнения скоростей слагаемые с N3, и т. д. Сумма всегда за-висит от N. Единственный случай, когда мгновенные скорости не зависят от разнообразия, - биологическое взаимодействие каждого таксона со все-ми другими. Этот факт является прямым следствием, во-первых, устанав-ливаемой эмпирически линейной зависимости N+(t), N-(t) на длительных интервалах геологического времени, и во-вторых, описания скоростей как зависящих от биотических отношений. Последнее вполне соответствует обычным эволюционным представлениям. Тем не менее, утверждение об облигатном влиянии всех таксонов друг на друга, являющемся условием наблюдаемого процесса, производит впечатление необычного и желатель-но получить более подробное описание его механизма. В этом отношении пока-что можно высказать только некоторые предварительные соображе-ния.
В современной биоте невозможно обнаружить всеобщее взаимовли-яние таксонов (популяций). Более того, условием устойчивости современ-ных экосистем является их ограниченная связность (May, 1973; Свирижев, Логофет, 1978). Под устойчивостью в экологии подразумевается сохране-ние в экосистеме постоянного видового состава. В палеонтологии наблю-даются постоянные изменения, то есть устойчивость в смысле теоретичес-кой экологии здесь не имеет места. На больших временах экосистемы не должны отвечать условию малой связности, она может быть большой и да-же полной. Условием этого является передача биологического взаимодей-ствия по цепям биотических связей за время, много меньшее изучаемого палеонтологими. Таким способом можно согласовать малую связность современных ненарушенных экосистем (она может быть более высокой при специальной организации связей) с требованием полной связности древних на больших временах, вытекающим из анализа скоростей.
Полная связность легче осуществляется в некоторых группах совме-стно обитающих таксонов. Тогда наблюдаемые скорости появления и вы-мирания есть суммы скоростей во всех таких группах. Прямо указать на какие-либо биологические совокупности, репрезентирующие описывае-мые гипотетические группы, затруднительно. Число групп на интервалах линейных зависимостей N+(t), N-(t) должно быть постоянным или меняться мало. Их соотношение с древними сообществами, описываемыми палеоэ-кологами, неизвестно. Представляет интерес эволюция сообществ. По име-ющимся данным (Bambach, 1986; Sepkoski et al., 1981), видовое богатство сообществ хорошо коррелировано с таксономическим разнообразием, ко-торое возрастает также за счет увеличения различий между сообществами (Sepkoski, 1988). Следовательно, число сообществ должно меняться мед-леннее разнообразия. Кроме сообществ, роль сильно связанных групп мо-гут играть и другие совокупности, например, население крупных биохо-рий, длительное сохранение числа которых в течение длительных времен-ных интервалов вполне вероятно.
Длительное существование многих сообществ животных характер-ного состава, фиксируемое палеонтологическими исследованиями, позво-ляет предположить устойчивое существование определенной структуры биологических отношений, которая обеспечивает в общем равномерный ход диверсификации без взрывоподобных явлений, разрушающих устой-чивость биоты. Симптоматично некоторое усиление различий между сооб-ществами в палеозое (Sepkoski, 1988). Структура связей внутри сообществ и между ними не может быть постоянно сохраняющейся и нарушается вследствие различных процессов, сочетающих эволюцию, миграцию и вы-мирание таксонов и сопряженные с ними изменения экосистем, в связи с чем могут возникать колебания скоростей различного масштаба вплоть до крупных. Может быть, но не является обязательным влияние внешних спу-сковых причин. Эта картина близка к представлениям о когерентной и не-когерентной эволюции и биоценотических кризисах (Жерихин, 1987; Ка-ландадзе, Раутиан, 1993).
В группах с высокой связностью для устойчивого развития таксоны должны быть как правило специализированными и коадаптированными. Поэтому циклы специализации-деспециализации ( Каландадзе, Раутиан, 1993) кажутся подходящим качественным описанием возможного меха-низма колебания скоростей.
При такой структуре биоты вполне естественной будет частая встре-чаемость эволюционных параллелизмов: сходные морфогенетические ог-раничения в эволюции близкородственных таксонов приводят к появле-нию однотипных рядов форм в сходных биоценотических структурах. Са-мо существование устойчивого набора структур такого типа, каждая из ко-торых препятствует дивергенции и способствует тенденции к преоблада-нию филетического вымирания, стабилизирует разнообразие биоты. Воз-можно, длительное, с ордовика по пермь сохранение палеозойского разно-образия морских животных обусловлено влиянием подобных факторов, а резкий рост разнообразия в ордовике сопряжен с перестройкой и увеличе-нием числа сообществ при формирования ярусности бентоса ( Ausich, Bot-tjer, 1985).
Линейность графиков N+(t), N-(t) и связанные с этим явления легче объяснить на основе представления о группах взаимосвязанных таксонов, но их существование не является необходимым. Биологические взаимо-влияния при наличии таких групп неизбежно передаются и между группа-ми, которые не могут быть сильно изолированными в масштабах геологи-ческого времени. Это делает всю биоту сильно связанной системой неза-висимо от большей или меньшей биологической роли групп. В развитии таких систем, как правило, возникают различные, в том числе широкомас-штабные колебания.
Представляет большой интерес совпадение основных минимумов разнообразия морских и различных континентальных животных. Оно мо-жет определяться высокой связностью всей биоты Земли. Современные представления о процессах, происходящих после внешнего воздействия на биоту (как хорошо документированный импакт на границе мела и палеоге-на), могут описывать, в сущности, поведение сильно связанных систем (ступенчатые вымирания - Алексеев, 1989; Барсков и др., 1996; Жерихин, Раутиан, 1999; колебания в компьютерных моделях связанных древних экосистем - Plotnick, McKinney, 1993). Переход от них к представлениям о возможности также автоколебаний большой амплитуды труден только психологически.
Вопрос об автоколебаниях биоразнообразия состоит в выборе между возможностью и необходимостью. Нет сомнений в возможности автоколе-баний в системах с высокой связностью, но должны ли они обязательно возникать в эволюции древней биоты зависит от ее конкретных характе-ристик. Геологическое время скорее всего достаточно велико для уверен-ной реализации этой возможности и ответ на поставленный вопрос ближе к “должны”, чем “могут”.
Определенный интерес может представлять диверсификация конти-нентальных тетрапод. Если дальнейшие исследования подтвердят пригод-ность логистической модели, это будет означать слабо зависимое от меж-таксонных биотических отношений развитие в начале процесса и дальней-шее его протекание при нарастающем влиянии этой зависимости. Это со-вершенно другой тип диверсификации, чем линейная, которая соответст-вует предельно высокой связности системы (равной 1), тогда как логисти-ческая диверсификация определяется низкой связностью (при нулевой связности развитие экспоненциально). Линейное и экспоненциальное развитие можно рассматривать как некоторые идеализации, напоминаю-щие группировки и сообщества в теоретической биоценологии (Алехин, 1986). Первому типу очень близко соответствуют траектории, реально на-блюдаемые в большинстве изученных случаев. Полного соответствия вто-рому типу не выявлено ни в одном случае, а близкий к нему логистичес-кий процесс у тетрапод сомнителен. Интерес представляют данные о воз-можных других типах кривых N+(t), N-(t). Пока-что кривые другого вида не выявлены, кроме кембрийской части кривых для морских животных (рис. 3). На рис. 24, б и 28, б показаны кривые N-(t) для пресноводных жи-вотных и насекомых, обнаруживающие замедление вымираний к совре-менности, слабое в первом случае и отчетливое во втором. Замедление мо-жет быть связано с влиянием знаний современной фауны, затрудняющим изучение диверсификации древних животных.
Описание диверсификации простыми зависимостями означает опре-деленного вида связь мгновенных скоростей появления и вымирания с раз-нообразием (или отсутствие такой связи). Параметры описывающих про-цесс зависимостей определяются свойствами усредненного таксона и ус-редненных межтаксонных отношений. Однотипное развитие на длитель-ных интервалах геологического времени означает постоянство парамет-ров, то есть сохранение на всем интервале средних свойств таксонов и отношений.
Изучение диверсификации организмов геологического прошлого яв-ляется неполным и несовершенным без анализа скоростей появления и вы-мирания. К сожалению, определение скоростей для небольших временных интервалов - веков и их частей, сильно затруднено недостаточной точно-стью имеющихся в настоящее время изотопно-геохронологических (гео-хронометрических) шкал. То же относится к применению шкал абсолют-ного возраста во многих других разделах палеонтологии и геологии в це-лом.
Положение в этой области недавно охарактеризовано А. Г. Рубле-вым (1996), который рассмотрел все предложенные к середине 90-х годов изотопно-геохронологические шкалы, кончая шкалой Г. Одэна (Odin, 1994). Удовлетворительной, согласно анализу Рублева, может считаться часть шкалы от позднего мела до современности. Более древние отложе-ния датированы недостаточно. При строгом подходе погрешность возраста отложений должна быть равна аналитической погрешности. Тогда пригод-ными для исследования признаются в основном только малоизмененные туфы и бентониты в отложениях, положение которых в общей геохроноло-гической шкале выяснено точно и надежно. При соблюдении этих требо-ваний большинство датировок, используемых как опорные для предложен-ных к настоящему времени шкал, приходится отвергнуть, после чего оста-ется слишком мало уверенно датируемых опорных точек. В послекембрий-ской и допозднемеловой части шкалы Рублев приводит только следующие датировки, которые он считает надежными: граница раннего и среднего ордовика - 473 (?) млн. лет, начало лландейло - 464, граница ордовика и силура - 440, начало лудлова - 421, граница среднего и позднего девона - 367, граница раннего и среднего триаса ³238, начало тоара - 180-190, гра-ница ранней и средней юры <185, граница юры и мела >137, начало валан-жина »135, граница раннего и позднего мела - 97. В проекте Международ-ной комиссии по стратиграфии, представленном на 31 Геологическом кон-грессе (2000), отсутствуют датировки от начала силура до конца девона, внутри среднего и позднего карбона, в поздней перми, от начала триаса до конца юры и внутри палеогена.
На рис. 39 показан график N+(t) для родов морских животных от на- чала сеномана до современности (возраст границ - по шкале Ф. Градштей-на и Дж. Огга; Gradstein, Ogg, 1996). График демонстрирует линейное на- копление появившихся родов, которое, таким образом, может быть ис-пользовано в качестве временной шкалы. Целесообразность этого опреде-
ляется двумя обстоятельствами. Точки графика не лежат точно на прямой,
<![if !vml]> <![endif]>
<![endif]>
Рис. 39. Появление родов морских животных в позднем мелу и кайнозое
а группируются около нее. Имеются отклонения, когда наблюдаемая ско-рость появления родов ниже средней (кампан), либо выше нее (ранний - средний эоцен, плиоцен - плейстоцен). Продолжительность, принятая про-порциональной числу появлений родов, в первом случае будет меньше, во втором - больше, чем в шкале Градштейна и Огга. Можно пренебречь от-клонениями скорости появления родов, считать скорость постоянной и тем самым заведомо огрубить получаемую временную шкалу. Тогда стано-вится актуальным вопрос о ее конкурентоспособности: получаемые таким способом оценки продолжительностей временных интервалов должны быть не хуже, чем в имеющихся шкалах. Второе обстоятельство более фундаментально: всегда желательно использовать методы, имеющие на-дежное теоретическое обоснование.
Качество получаемой шкалы может быть оценено сравнением про-должительностей с таковыми в имеющихся геохронометрических изотоп-ных шкалах для общепризнанно считающейся наиболее точной и досто-верной позднемеловой - кайнозойской их части (табл. 4,5). Общая продол-жительность всего изученного интервала принята по Градштейну и Оггу в 98.9 млн. лет.
Для обобщенного сравнения шкал (табл. 5) могут быть использова-ны среднее и максимальное относительные отклонения продолжительнос-тей от их среднего арифметического в двух сравниваемых шкалах (чтобы не отдавать предпочтения ни одной из них).
Шкала, основанная на палеонтологических данных, по согласован-ности с изотопными шкалами в среднем несколько уступает их согласо-ванности между собой. Максимальные отклонения (39 и 52%) относятся к
Таблица 4
Продолжительности веков позднего мела - кайнозоя в разных шкалах
Число Длительность в млн. лет
появившихся I II III IV
родов
Сеноман 779 7.1 6.5 4.0 5.4
Турон 360 3.3 2.5 4.0 4.5
Коньяк 256 2.3 1.0 1.0 3.2
Сантон 356 3.3 3.5 4.0 2.3
Кампан 768 7.1 9.5 11.0 12.2
Маастрихт 650 6.0 8.1 7.0 6.3
Даний 357 3.3 2.8 6.0 4.0
Палеоцен 607 5.6 5.8 6.0 6.2
Ипр 830 7.5 5.8 7.0 5.8
Лютет 1185 10.8 8.4 6.0 7.7
Бартон+приабон 576 5.3 7.0 6.3 7.6
Рюпель 510 4.7 6.6 5.7 5.2
Хатт 438 4.0 6.3 5.0 4.7
Ранний миоцен 962 8.8 7.1 7.2 7.4
Средний миоцен 611 5.6 5.4 4.8 5.2
Поздний миоцен 632 5.8 5.9 5.7 5.9
Плиоцен 660 6.0 3.7 3.5 3.5
Плейстоцен 264 2.4 1.6 1.8 1.8
I - палеонтологические данные; II - шкала Американского геологического общества, 1990, использовавшаяся в предыдущих разделах этой работы; III - Одэн, 1994; IV - Градштейн и Огг, 1996
Таблица 5
Сравнение шкал позднего мела – кайнозоя
I/II I/III I/IV II/III II/IV III/IV
Среднее,% 13 15 12 10 11 10
Максимальное,% 39(24) 39(29) 26 36 52(29) 52(27)
коньяку, дительность которого в шкалах I и II принята 1 млн. лет и явно занижена. Максимальные отклонения без учета коньяка приведены в скоб-ках. Нетрудно получить более точное совпадение с изотопными шкалами, если на рис. 39 выделить более кратковременные прямолинейные участки и использовать по две опорные датировки для каждого из них. Здесь пред-намеренно взята единственная датировка (граница раннего и позднего ме-ла), так что шкала сопоставимая по качеству с имеющимися изотопными шкалами получена сразу для интервала в 100 млн. лет.
Практическая ценность изложенного метода может быть более ощу-тимой для допозднемеловой части геохронологической шкалы. Относи-тельные отклонения продолжительностей веков в трех сравниваемых изо-топных шкалах на интервале тремадок - альб (без поздней перми и раннего триаса) следующие:
II/III II/IV III/IV
Среднее,% 15 16 18
Максимальное,% 54 53 56
Они не лучше сравнительных показателей палеонтологической шка-лы для позднего мела - кайнозоя в сопоставлении ее с изотопными шкала-ми. Поэтому палеонтологическая геохронометрия может использоваться для общей ориентировки в допозднемеловой части шкалы. Для этого на графике N+(t), построенном с использованием любой изотопно-геохроно-метрической шкалы (или нескольких), выделяются прямолинейные интер-валы. В их пределах продолжительности веков и их частей определяются пропорционально установленному числу появлений родов. Для каждого интервала необходимы минимум две опорные изотопные датировки, жела-тельно близ его верхней и нижней границ. При наличии большего коли-чества опорных датировок скорость определяется методом наименьших квадратов.
В разделе “Кривые разнообразия” приведен график N+(t) по родам морских животных для всего фанерозоя и выделены три допозднемеловых более или менее прямолинейных интервала: арениг - турне, визе - ленерд и анизий - альб (рис. 4, стр. 17). Для этих интервалов здесь приводится дета-лизированная (где это было возможно) палеонтологическая шкала продол-жительностей (табл. 6; DN+ - число появившихся родов, t - продолжитель-ность в млн. лет, t - возраст нижней границы, t* - то же по Градштейну и Оггу). Опорные датировки отмечены звездочкой; первый десятичный знак приведен для удобства сравнения, он не может считаться значащим.
Таблица 6
Интерполяция геологического возраста по палеонтологическим данным для палеозоя и мезозоя
DN+ t t t*
Ранний арениг 340 4.7 485.0 *485.0
Поздний арениг 542 7.5 480.3
Ранний лланвирн 271 3.7 472.9 470.0
Поздний лланвирн 241 3.3 469.2
Ранний лландейло 260 3.6 465.9 464.0
Поздний лландейло 283 3.9 462.3
Ранний карадок 502 6.9 458.4 458.0
Средний карадок 396 5.4 451.5
Поздний карадок 195 2.7 446.1
Ранний ашгилл 198 2.7 443.4 449.0
Средний ашгилл 352 4.8 440.7
Поздний ашгилл 75 1.0 435.9
Ранний лландовери 111 1.5 434.8 443.0
Средний лландовери 204 2.8 433.3
Поздний лландовери 303 4.2 430.5
Ранний венлок 396 5.4 426.4 428.0
Поздний венлок 339 4.7 420.9
Ранний лудлов 295 4.0 416.3 423.0
Поздний лудлов 257 3.5 412.2
Пржидол 231 3.2 408.7 419.0
Жедин 459 6.3 405.5 417.0
Зиген 496 6.8 399.2
Ранний эмс 422 5.8 392.4 400.0
Поздний эмс 334 4.6 386.6
Эйфель 607 8.3 382.0 391.0
Ранний живет 217 3.0 373.7 380.0
Поздний живет 262 3.6 370.7
Ранний фран 181 2.5 367.1 370.0
Средний фран 122 1.7 364.6
Поздний фран 185 2.5 363.0
Ранний фамен 118 1.6 360.4 364.0
Средний фамен 224 3.1 358.8
Поздний фамен 256 3.5 355.7
Ранний турне 393 5.4 352.2 354.0
Поздний турне 353 4.8 346.8
Ранний визе 337 9.8 342.0 *342.0
Поздний визе 513 14.8 332.2
Ранний серпухов 259 7.6 317.3 327.0
Поздний серпухов 41 1.2 309.7
Ранний башкир 136 4.0 308.5 323.0
Поздний башкир 141 4.1 304.5
Раннемосковский 102 3.0 300.4 311.0
Позднемосковский 243 7.1 297.5
Касимовский 124 3.6 290.4 303.0
Гжельский 101 2.9 286.8 296.5
Ассельский 198 5.8 283.8 290.0
Сакмарский 246 7.2 278.1 282.0
Ранний ленерд 75 2.2 270.9 269.0
Средний ленерд 194 5.7 268.7
Поздний ленерд 241 7.0 263.0
Гвадалупий - - 256.0 *256.0
Ранний анизий 117 2.5 241.7 *241.7
Средний анизий 133 2.8 239.2
Поздний анизий 133 2.8 236.4
Ранний ладин 104 2.2 233.6 234.3
Поздний ладин 288 6.2 231.4
Ранний карний 193 4.1 225.4 227.4
Поздний карний 239 5.0 221.3
Ранний норий 173 3.6 216.3 220.7
Средний норий 164 3.4 212.6
Поздний норий 82 1.7 209.2
Рэт 105 2.2 207.5 209.6
Геттанг 151 3.2 205.3 205.7
Ранний синемюр 111 2.3 202.1 201.9
Поздний синемюр 153 3.2 199.8
Ранний плинсбах 144 3.0 196.5 195.3
Поздний плинсбах 165 3.5 193.5
Ранний тоар 172 3.6 190.0 189.6
Поздний тоар 93 2.0 186.4
Аален 150 3.2 184.5 180.1
Ранний байос 167 3.5 181.3 176.5
Поздний байос 246 5.2 177.8
Ранний бат 119 2.5 172.6 169.2
Средний бат 68 1.4 170.1
Поздний бат 161 3.4 168.7
Ранний келловей 109 2.3 165.3 164.4
Средний келловей 102 2.2 163.0
Поздний келловей 99 2.1 160.9
Ранний оксфорд 203 4.3 158.8 159.4
Средний оксфорд 46 1.0 154.5
Поздний оксфорд 213 4.5 153.5
Ранний кимеридж 141 3.0 149.1 154.1
Поздний кимеридж 120 2.5 146.1
Ранний титон 194 4.1 143.6 150.7
Поздний титон 216 4.6 139.5
Берриас 128 2.7 134.9 144.2
Валанжин 250 5.3 132.2 137.0
Ранний готерив 113 2.4 126.9 132.0
Поздний готерив 75 1.6 124.6
Баррем 224 4.7 123.0 127.0
Ранний апт 162 3.4 118.3 121.0
Поздний апт 196 4.1 114.9
Ранний альб 161 3.4 110.7 112.2
Средний альб 101 2.1 107.4
Поздний альб 301 6.4 105.2
Сеноман - - 98.9 *98.9
Шкала приведена здесь не для выяснения “истинных” датировок и продолжительностей, а для демонстрации возможности использования па-леонтологических данных. Возраст границ, как и следовало ожидать, то больше, то меньше отличается от указываемого в изотопно- геохрономет-рических шкалах, но примерно так же, как он различается в разных изо-топных шкалах. Это может быть проиллюстрировано числом границ, воз-раст которых в разных шкалах по абсолютной величине отличается на D (табл. 7; с аренига по альб без поздней перми и раннего триаса):
Продолжительности веков при сравнении со шкалой Градштейна и Огга характеризуются средним относительным отклонением 17% (от сред- него арифметического, как и выше) и максимальным отклонением 46% (берриас). Эти же величины описывают в целом изменения наблюдаемой скорости появления родов и могут служить для общей оценки точности получаемых по палеонтологическим данным шкал. В условиях неопреде-
Таблица 7
Сравнение шкал палеозоя и мезозоя
I/IV II/III II/IV III/IV
15£D<20 0 0 0 0
10£D<15 7 6 2 2
5£D<10 11 23 13 21
3£D<5 8 6 13 8
1£D<3 11 6 14 12
D<1 11 4 6 2
ленности оценка искомой продолжительности веков как среднего арифме-тического по двум шкалам формально может считаться наилучшей из воз-можных, но приходится иметь ввиду, во-первых, принятое допущение о постоянстве скорости появления родов, и во-вторых, недостаточную в большинстве случаев обоснованность продолжительностей веков в изото-пных временных шкалах.
Все приведенные выше количественные показатели сравнения раз-ных временных шкал характеризуют преимущественно их взаимное соот-ветствие, но не обязательно точность. Связь этих показателей вполне оче-видна: чем выше точность шкал, тем они больше соответствуют друг дру-гу, но разные шкалы не являются независимыми, так как совокупность конкретных опорных датировок, лежащая в их основе, значительно пере-крывается. Поэтому точность шкал может не вполне соответствовать их взаимной согласованности (если не являются достаточно надежными слишком многие датировки). Проявляя осторожность, кажется возможным оценить общую точность палеонтологической шкалы, удвоив приведенные выше оценки, то есть 30-40% для средней относительной точности и около 100% для максимальных отклонений. Эта общая оценка точности палеон-тологической интерполяции в целом соответствует колебаниям наблюдае-мой скорости появления родов на арениг-турнейском, визе-ленердском и анизий-альбском интервалах, показанным на рис. 7.
Усредненная оценка точности палеонтологической геохронометрии является вполне удовлетворительной в условиях, когда на продолжитель-ных интервалах геологического времени отсутствуют изотопные датиров-ки границ, признаваемые всеми исследователями. Это, однако, не дает воз-можности оценить погрешность того или иного конкретного временого интервала, продолжительность которого установлена путем интерполяции по палеонтологическим данным.
Погрешность конкретного определения в принципе может быть ука-зана только для изотопных датировок, получаемых на основании физичес-кого закона, что определяет ведущее (теоретически) значение изотопного датирования при вспомогательной роли других методов. Все же интерпо-ляция по палеонтологическим данным может оказаться полезной. В приве-денном выше демонстративном варианте шкалы пять опорных датировок дополнены 89 интерполированными и существенно повышена детальность шкалы. Применение здесь линейной интерполяции определяется видом эмпирической кривой время - число появившихся родов. В случае кривых другого вида (кембрий) интерполяция может проводиться на основе дру-гих зависимостей, если материал позволяет выяснить их вид. Основной во-прос в дальнейшем обосновании и развитии этого метода - выяснение ре-альных изменений скорости появления.
Наблюдаемые изменения скоростей появления родов зависят от нес-кольких факторов. При постоянной в действительности скорости появле-ния нового разнообразия наблюдаемая скорость появления родов будет уменьшаться или увеличиваться, если продолжительность века в исполь-зуемой изотопно-геохронометрической шкале соответственно преувеличе-на или преуменьшена. Имеют значение также возможные различия страти-графических корреляций, которые использованы для построения изотоп-ной шкалы и для обобщения палеонтологических данных. Кроме того, не-равномерная палеонтологическая изученность отложений определяет вы-теснение фиксируемых палеонтологическими материалами появлений ро-дов из менее изученных подразделений в более молодые лучше изучен-ные. Наблюдаемая скорость появления родов будет преуменьшена по сра-внению с действительной в плохо изученных веках и преувеличена в сле-дующих за ними лучше изученных (о контроле палеонтологической изу-ченности и возможностях компенсации связанных с ней дефектов см. гла-ву I).
Дополнительные искажения могут возникать вследствие несовер-шенства системы на уровне родов и дисперсии пуассоновского процесса появлений. Дисперсия равна ожидаемому числу появлений. Этот фактор приобретает большее значение, когда ожидаемое число появлений мало. При ожидаемом появлении 300-500 родов среднее квадратичное отклоне-ние 17-22 рода. Относительная погрешность, соответствующая одному среднему квадратичному отклонению, составит 4-6%. Если ожидаемое число появлений 50-100, эта погрешность возрастает до 10-14%, что огра-ничивает возможности измерения коротких временных интервалов. Для измерения продолжительности фазы (время формирования хронозоны) чи-сла появляющихся родов может оказаться недостаточно. Переход к работе с числом видов может скомпенсировать эту потерю точности, но на уровне видов может быть велико влияние погрешностей, обусловленных неравно-мерной палеонтологической изученностью.
Наибольший интерес представляют действительные изменения ско-рости появления нового разнообразия. Пути эмпирического исследования этого вопроса вполне очевидны: повышение детальности и точности изо-топной временной шкалы и полноты палеонтологической изученности. Теоретическое исследование подразумевает математическое описание ко-лебаний скорости появления в составе более общей теории диверсифика-ции. Описание такого типа может дать более точные обобщенные характе-ристики возможных колебаний скорости появления, но вряд ли позволит оценить погрешности конкретных продолжительностей.
Статус палеонтологической геохронометрии определяется как вспо-могательный ее зависимостью от некоторого исходного допущения о по-ведении скорости появления, а не связью с точно установленным законом природы. В этом палеонтологическая геохронометрическая шкала анало-гична известной шкале А. Холмса (1963), которая была предложена в 1947 году и основана на предположении о постоянстве максимальной скорости осадконакопления в геологической истории. Холмс использовал пять опорных радиоизотопных дат и на этой основе интерполировал продолжи-тельности периодов по максимальной суммарной мощности их отложе-ний. Шкала была очень популярна и широко использовалась в течение 20-25 лет в 50-е годы прошлого века и позже (в уточненном варианте 1960 г., когда она была пересмотрена и дополнена новыми данными). По сравне-нию с современной шкалой Градштейна и Огга относительное отклонение продолжительностей периодов в последнем варианте шкалы Холмса в среднем составило 27%, достигая 54% для силура и 100% для кембрия. Это не явилось препятствием к использованию шкалы Холмса для многих исследований и, в частности, крупных обобщений по геохимии осадочно-го процесса (Ронов, 1964; Гаррелс, Маккензи, 1974), полностью сохранив-ших свое значение.
Основное допущение хронометрии по палеонтологическим данным при линейной интерполяции состоит в постоянной скорости появления ро-дов, поэтому формируемая в результате временная шкала непригодна для изучения этой скорости в совокупности групп, использованной для ее пос-троения. Это не исключает анализа скорости появления в отдельных груп-пах организмов, составляющих сравнительно небольшую часть всей сово-купности.
1. Графики время - мгновенное число таксонов для родов, семейств и отрядов морских животных сходны в положении основных минимумов и максимумов при различиях общей конфигурации. Связь числа таксонов смежных рангов приблизительно описывается эмпирической зависи-мостью
Ni+1 = A(lnNi) r,
где Ni - число таксонов, Ni+1 - число таксонов следующего более высокого ранга, A, r - параметры.
2. На графиках время - накопленное число появившихся таксонов, время - накопленное число вымерших таксонов для родов и семейств мор-ских животных выявляется зависимость, близкая к линейной на протяжен-ных интервалах геологического времени: арениг - турне, визе - ленерд, анизий - альб, кайнозой. На этих интервалах скорости появления и выми-рания родов и семейств вариируют около некоторых постоянных. Зависи-мость скоростей появления и вымирания от числа таксонов отсутствует. То же наблюдается у пресноводных животных и насекомых. Отношение средней скорости появления и вымирания морских животных на указан-ных временных интервалах к средней скорости в фанерозое не зависит от ранга таксонов (семейств или родов, кроме кайнозоя).
3. Независимость скоростей от разнообразия свидетельствует о су-ществовании биотической саморегуляции скоростей, которая может со-провождаться возникновением автоколебаний, что следует учитывать при интерпретации минимумов кривой разнообразия. Некоторые минимумы разнообразия, характеризуемые в литературе как “массовые вымирания”, не обусловлены сколько-нибудь существенным увеличением скорости вы-мирания, а связаны с уменьшением скорости появления. Следует избегать применения термина “массовое вымирание” к недостаточно изученным минимумам разнообразия.
4. Сопоставление кривых разнообразия морских и пресноводных жи-вотных, а также континентальных тетрапод и насекомых показывает сов-падение основных минимумов разнообразия.
5. Сравнение кривых числа семейств насекомых по двум независимо составленным спискам (Лаб. артропод ПИН и К. Лабандейра, 1994), силь-но различающимся как по составу семейств, так и по датировкам, показы-вает высокое сходство кривых (коэффициент корреляции 0.986). Различия на некоторых участках каждый раз связаны с разными датировками круп-ных местонахождений.
6. Возможны две независимые (но совместимые) интерпретации вы-сокой стабильности кривых разнообразия в процессе изучения: а) количес-твенная - кривые отражают результат опосредованного соизмерения мор-фологического разнообразия в разные моменты времени; б) качественная - форма кривых обусловлена коллективным поведением групп. Количест-венная интерпретация дает больше возможностей для развития метода и различных приложений.
7. Морфологическое разнообразие может измеряться непосредствен-но, как сумма таксономических расстояний внутри изучаемой совокупнос-ти организмов (с минимальным использованием системы). Это теорети-ческий предел детальности исследований по диверсификации ископаемых организмов.
8. Погрешности измерения морфологического разнообразия обусло-влены неравномерной и неоднородной палеонтологической изученностью крупных групп и отложений разного возраста, измерением разнообразия числом таксонов, погрешностями стратиграфической корреляции и изо-топно-геохронологической шкалы. Следствием палеонтолого-стратигра-фических погрешностей может быть отсутствие статистической репрезен-тативности изученной части палеонтологической летописи по отношению к биоразнообразию геологического прошлого. Это исключает применение любых количественных характеристик кратковременных явлений без ре-конструкции генеральной совокупности. Следует использовать оценки разнообразия при изученности, приближающейся к 100%.
9. Актуальное (наблюдаемое, опубликованное) разнообразие и по-тенциальное (доступное для полноценного изучения) разнообразие, выде-ленные ранее Н. Ньюеллом (Newell, 1959), следует дополнить еще двумя категориями - полное (захоронившееся) и общее (существовавшее) разно-образия. Основным фактором, искажающим палеонтолого-стратиграфи-ческие данные, может быть неравномерная представленность полного раз-нообразия в потенциальном. Предлагается реконструировать полное раз-нообразие последовательным суммированием потенциального разнообра-зия отдельных территорий (изученность которого должна каждый раз кон-тролироваться) с дальнейшей экстраполяцией на всю площадь обитания древних фаун.
10. Анализ данных по морским гастроподам кампана - монса с целью выяснения полноты актуального разнообразия по отношению к потенци-альному показывает возможную высокую изученность гастропод кампана.
11. Появление родов морских животных можно использовать для де-тализации изотопно-геохронологической шкалы.
Алексеев А. С. 1984. Количественный анализ вымирания на рубеже мезозоя и кайнозоя // Бюл. Моск. о-ва испыт. природы. Отд. геол. Т. 59. № 2. С. 87-109.
Алексеев А. С. 1986. Вымирание на рубеже мезозоя и кайнозоя // Природа. № 1. С. 57-60.
Алексеев А. С. 1989. Глобальные биотические кризисы и массовые вымирания в фанерозойской истории Земли // Биотические события на ос-новных рубежах фанерозоя. М.: МГУ. С. 22-47.
Алексеев А. С. 1989. Массовые вымирания и их место в развитии биосферы // Осадочная оболочка Земли в пространстве и времени. Страти-графия и палеонтология. Докл. сов. геологов на 28-й сесс. Междунар. геол. конгр. М.: Наука. С. 27-34.
Алексеев А. С., Дмитриев В. Ю., Пономаренко А. Г. 2001. Эволюция таксономического разнообразия. М.: ГЕОС. 125 с.
Алехин В. В. 1986. Теоретические проблемы фитоценологии и степе-ведения. М.: МГУ. 211 с.
Амитров О. В., Бланк М. Я., Горбач Л. П., Джабарова В. В., Калише-вич Т. Г., Макаренко Д. Е., Мартинсон Г. Г., Миронова Л. В., Мороз С. А., Толстикова Н. В., Яркин В. И. 1981. Класс Gastropoda. Брюхоногие мол-люски // Развитие и смена моллюсков на рубеже мезозоя и кайнозоя. М.: Наука. С. 72-123.
Барсков И. С., Богословская М. Ф., Кузина Л. Ф., Журавлева Ф. А., Шиманский В. Н., Яцков С. В. 1994. Динамика изменения родового разно-образия и экологической структуры цефалопод (кембрий - карбон) // А. Ю. Розанов, М. А. Семихатов (ред.). Экосистемные перестройки и эволюция биосферы. Вып.1. М.: Недра. С. 86-94.
Барсков И. С., Жерихин В. В., Раутиан А. С. 1996. Проблемы эволю-ции биологического разнообразия // Журн. общей биол. T. 57, № 2. С. 14-39.
Беккер-Мигдисова Е. Э. 1962. Некоторые новые полужесткокрылые и сеноеды // Палеонтол. журн. № 1. С. 89-104.
Беклемишев В. Н. 1994. Методология систематики. М.: KMK Scienti-fic Press. 250 с.
Виноградова Н. Г. 1977. Фауна шельфа, материкового склона и абис-сали // Океанология. Биология океана. Т. 1. Биологическая структура океа-на. М.: Наука. С. 178-198.
Гаррелс Р., Маккензи Ф. 1974. Эволюция осадочных пород. М.: Мир. 160 с.
Дмитриев В. Ю. 1978. Некоторые вопросы изучения изменений сис-тематического разнообразия ископаемых организмов // Палеонтол. журн. № 2. С. 122-132.
Дмитpиeв B. Ю. 1989. O дивepcификaции мopcкиx живoтныx фaнe-poзoя // Ocaдoчнaя oбoлoчкa Зeмли в пpocтpaнcтвe и вpeмeни. Cтpaтигpa-фия и пaлeoнтoлoгия. Докл. сов. геол. на 28-й сесс. Междунар. геол. конгр. M.: Hayкa. C. 6-13.
Дмитpиeв B. Ю. 2001. Кривые разнообразия. Палеонтологическая интерполяция в геохронометрии // Алексеев А. С., Дмитриев В. Ю., Поно-маренко А. Г. Эволюция таксономического разнообразия. М.: ГЕОС. С. 19-67, 111-118.
Дмитриев В. Ю., Пономаренко А. Г., 2001. Эволюция разнообразия континентальных биот // Там же. С. 68-95.
Дмитpиeв B. Ю., Жерихин В. В. 1988. Изменения разнообразия се-мейств насекомых по данным метода накопленных появлений // Меловой биоценотический кризис и эволюция насекомых. М.: Наука. С. 208-215.
Дмитриев В. Ю., Пономаренко А. Г., Расницын А. П. 1994а. Разнооб-разие организмов геологического прошлого. Состояние проблемы // В. Е. Соколов, Ю.С. Решетников (ред.). Биоразнообразие. Степень таксономи-ческой изученности. М.: Наука. С. 12-19.
Дмитриев В. Ю., Пономаренко А. Г., Расницын А. П. 1994б. Дина-мика таксономического разнообразия неморской биоты // А. Ю. Розанов, М. А. Семихатов (ред.). Экосистемные перестройки и эволюция биосферы. Вып.1. М.: Недра. С. 167-174.
Дмитриев В. Ю., Пономаренко А. Г., Расницын А. П. 1995. Динами-ка таксономического разнообразия неморской водной биоты // Палеонтол. журн. № 4. С. 3-9.
Жерихин В. В. 1978. Развитие и смена меловых и кайнозойских фау-нистических комплексов. М.: Наука. 198 с. (Тр. Палеонтол. ин-та АН СССР; Т. 165).
Жерихин В. В. 1987. Биоценотическая регуляция эволюции // Пале-онтол. журн. № 1. С. 3-12.
Жерихин В. В., Раутиан А. С. 1999. Кризисы в биологической эволю-ции // Анатомия кризисов. М.: Наука. С. 29-47.
Каландадзе Н. Н., Раутиан А. С. 1993. Юрский экологический кризис сообщества наземных тетрапод и эвристическая модель сопряженной эво-люции сообщества и биоты // Проблемы доантропогенной эволюции био-сферы. М.: Наука. С. 60-95.
Калугинa Н. С. 1980. Насекомые в водных экосистемах прошлого // Родендорф Б. Б., Расницын А. П. (ред.). Историческое развитие класса на-секомых. М.: Hаука. С. 145-148. (Тр. Палеонтол. ин-та АH СССР; Т. 178).
Красилов В. А. 1992. Охрана природы: принципы, проблемы, прио-ритеты. М.: Внииприрода. 173 с.
Левен Э. Я. 1970. О происхождении высших фузулинид // Палеон-тол. журн. № 3. С. 18-25.
Марков А. В. 2001. О причинах корреляции скоростей появления и вымирания родов в фанерозойской морской биоте // Розанов А. Ю., Поно-маренко А. Г., Федонкин М. А. (ред.) . Экосистемные перестройки и эво-люция биосферы. Вып. 4. М.: ПИН. С. 87-96.
Марков А. В. 2002. О механизмах роста таксономического разнооб-разия морской биоты в фанерозое // Палеонтол. журн. № 2. С. 3-13.
Марков А. В., Наймарк Е. Б. 1994. Линии выживания как доказатель-ство реальности высших таксонов // Палеонтол. журн. № 2. С. 3-11.
Нейман А. А., Зезина О. Н., Семенов В. Н. 1977. Донная фауна шель-фа и материкового склона // Океанология. Биология океана. Т. 1. Биологи-ческая структура океана. М.: Наука. С. 269-281.
Песенко Ю. А. 1982. Принципы и методы количественного анализа в фаунистических исследованиях. М.: Наука. 287 с.
Пономаренко А. Г. 1990. Жуки. Scarabaeida // Расницын А.П. (ред.).
Позднемезозойские насекомые Восточного Забайкалья. М.: Наука. С. 39-88. (Тр. Палеонтол. ин-та АН СССР; Т. 239).
Пономаренко А. Г., Дмитриев В. Ю. 1993. Эволюция разнообразия и устойчивость экосистем // А. Ю. Розанов (ред.). Проблемы доантропоген-ной эволюции биосферы. М. Наука. С. 54-59.
Пономаренко А. Г., Дмитриев В. Ю. 2001. Эволюция разнообразия континентальной биоты // Биоразнообразие в истории Земли. Тез. докл. XLVII сесс. Палеонтол. общ. СПб.: ВСЕГЕИ. С. 83-84.
Пузаченко Ю. Г., Дроздова Н. Н. 1986. Площадь охраняемых терри-торий // Итоги и перспективы заповедного дела в СССР. М.: Наука. С. 72-109.
Развитие и смена моллюсков на рубеже мезозоя и кайнозоя. 1981. М.: Наука. 142 с.
Расницын А. П. 1988. Проблема глобального кризиса наземных био-ценозов в середине мелового периода // Пономаренко А. Г. (ред.). Мело-вой биоценотический кризис и эволюция насекомых. М.: Наука. С. 191-207.
Расницын А. П. 1989. Динамика семейств насекомых и проблема ме-лового биоценотического кризиса // Ocaдoчнaя oбoлoчкa Зeмли в пpocт-paнcтвe и вpeмeни. Cтpaтигpaфия и пaлeoнтoлoгия. Докл. сов. геол. на 28-й сесс. Междунар. геол. конгр. M.: Hayкa. C. 35-40.
Розанов А. Ю. 1973. Закономерности морфологической эволюции археоциат и вопросы ярусного расчленения нижнего кембрия. М.: Наука. 165 с. (Тр. Геол. ин-та АН СССР; Вып. 241).
Ронов А. Б. 1964. Общие тенденции в эволюции состава земной ко-ры, океана и атмосферы // Геохимия. № 8. С. 715-743.
Рублев А. Г. 1996. Современное состояние изотопно-геохронологи-ческой шкалы фанерозоя // Стратиграфия, геологическая корреляция. М.: Наука. Т. 4. № 6. С. 3-13.
Свирижев Ю. М., Логофет Д. О. 1978. Устойчивость биологических сообществ. М.: Наука. 352с.
Холмс А. 1963. Пересмотренная геохронологическая шкала // Геохи-мия литогенеза. М.: Иностранная литература. С. 390-430.
Чайковский Ю. В. 1990. Элементы эволюционной диатропики. М.: Наука. 271 с.
Шевырев А. А. 1990. Аммоноидеи и хроностратиграфия триаса. М. Наука. 179 с. (Тр. Палеонтол. ин-та АН СССР; Т. 241).
Шиманский В. Н., Соловьев А. Н. 1982. Рубеж мезозоя и кайнозоя в развитии органического мира. М.: Наука. 39 с.
Ausich W. I., Bottjer D. J. 1985. Phanerozoic tiering in suspension-feed-ing communities on soft substrata: implications for diversity // Valentine J. W. (ed.). Phanerozoic diversity patterns: Profiles in macroevolution. Princeton: Princeton Univ. Рress. P. 255-274.
Bambach R. K. 1986. Phanerozoic marine communities // Raup D. M., Jablonski D. (eds). Patterns and processes in the history of life. Berlin: Sprin-ger. P. 407-428.
Benton M. J. 1985. Mass extinction among families of none-marine tetra-pods // Nature. Vol. 306. № 6031. P. 811-814.
Benton M. J. 1987. Mass extinction among families of none-marine tetra-pods: The data // Mem. Soc. geol France. № 150. P. 21-32.
Benton M. J. 1989. Patterns of evolution and extinction in vertebrates // Evolution and the fossil record. London: Belhaven Press. P. 218-241.
Benton M. J. (ed.). 1993. The fossil record (2nd edition). London: Chap-man and Hall. 839 р.
Benton M. J. 1994. Late Triassic to Middle Jurassic extinctions among continental tetrapods: testing the pattern // Fraser N. C., Sues H.-D. (eds). In the shadow of the dinosauirs. Cambridge: Cambridge Univ. Press. P. 366-397.
Benton M. J. 1995. Diversification and extinction in the history of life // Science. Vol. 268, № 5207. P. 52-58.
Connor E. F., McCoy E. D. 1979. The statistics and biology of the species - area relationship // Amer. Natur. Vol. 113. № 6. P. 791-833.
Courtillot V., Gaudemer Y. 1996. Effects of mass extinctions on biodi-versity // Nature. Vol. 381. № 6578. P. 146-148.
Dmitriev V. Yu., Ponomarenko A. G., 2002. Dymamics of insect taxono-mic diversity // Rasnitsyn A. P., Quicke D. L. J. (eds). History of insects. Dord-recht: Kluver Acad. Publ. P. 325-331.
Doyle J. A. 1993. Russian Paleoentomology // Science. Vol. 262. № 5133. P. 492.
Fenster E. J., Hecht M. K., Sorhannus U. 1991. Problems in the measure-ment of morphological rates of change // Ann. zool. fenn. Vol. 28. № 3-4. P. 165-174.
Flessa K. W., Jablonski D. 1985. Declining Phanerozoic background ex-tinction rates: effect of taxonomic structure ? // Nature. Vol. 113, № 5999. P. 216-218.
Foote M. 1993. Discordance and concordance between morphological and taxonomic diversity // Paleobiology. Vol. 19. № 2. P. 185-204.
Foote M. 2000. Origination and extinction components of taxonomic di-versity: general problems // Erwin D. H., Wing S. L. (eds). Deep time. Paleobio-logy’s perspective. Lawrence: Allen Press. P. 74-102. (Suppl. to Paleobiology. Vol. 26. № 4).
Foote M., Sepkoski J. J., Jr. 1999. Absolute measures of the completeness of the fossil record // Nature. Vol. 398. № 6726. P. 415-417.
Gradstein F. M., Ogg J. 1996. A Phanerozoic time scale // Episodes. Vol. 19. № 1-2. P. 3-5.
Graffin G. 1992. A new locality of fossiliferous harding sandstone: Evi-dence for freshwater Ordovician vertebrates // J. vertebr. Paleontol. Vol. 12. № 1. P. 1-10.
Gray J. 1988. Evolution of the freshwater ecosystem: the fossil record // Palaeogeogr., Palaeoclimatol., Palaeoecol. Vol. 62. 214 p.
Grimaldi D. A. (ed.). 1990. Insects from the Santana formation, Lower Cretaceous, of Brasil // Bull. Amer. Museum Natur. History. № 195. 191 p.
Harper C. W. 1975. Standing diversity of fossil groups in succesive inter-vals of geologic time: a new measure // J. Paleontol. Vol. 49. № 4. P. 752-757.
Hoffman A. 1985. Biotic diversification in the Phanerozoic: diversity in-dependence // Palaeontology. Vol. 28. № 2. P. 387-391.
Holland S. M. 2001. The quality of the fossil record: a sequence stratigra-phic perspective // Erwin D. H., Wing S. L. (eds). Deep time. Paleobiology’s perspective. Lawrence: Allen Press. P. 148-168. (Suppl. to Paleobiology. Vol. 26. № 4).
Jarzembowski E., Ross A. 1993. Time flies: the geological record of in-sects // Geology today. Nov.-dec. P. 218-223.
Jell P. A., Roberts J. (eds). 1986. Plant and invertebrates from the Lower Cretaceous Koonwarra fossil bed, South Gippsland, Victoria // Assoc. Australa-sian Paleontol. Mem. 3. 205 p.
Kirchner J. W., Weil A. 1998. No fractals in fossil extinction statistics // Nature. Vol. 395. № 6700 P.337-338.
Koch C. F. 1978. Bias in the published fossil record // Paleobiology. Vol. 4. № 3. P. 367-372.
Koch C. F. 1987. Prediction of sample size effects on the measured tem-poral and geographic distribution patterns of species // Paleobiology. Vol. 13. № 1. P. 100-107.
Koch C. F., Morgan J. P. 1988. On the expected distribution of species ranges // Paleobiology. Vol. 14. № 2. P. 126-138.
Labandeira C. C. 1994. A compendium of fossil insects families // Con-trib. in biol. and geol. Milwaukee Public Museum. № 88. 71 p.
Labandeira C. C., Sepkoski J. J., Jr. 1993. Insect diversity in the fossil re-cord // Science. Vol. 261. № 5119. P. 310-315.
Lasker H. R. 1976. Effects of differential preservation on the measure-ment of taxonomic diversity // Paleobiology. Vol. 2. № 2. P. 84-93.
Lasker H. R. 1978. The measurement of taxonomic evolution: preserva-tional consequences // Paleobiology. Vol. 4. № 2. P. 135-149.
Maxwell W. D., Benton M. J. 1990. Historical tests of the absolute comp-letness of the fossil record of tetrapods // Paleobiology. Vol. 16. № 3. P. 322-335.
May R. M. 1973. Stability and сomplexity in Model Ecosystems. Prince-ton: Princeton Univ. Press. 285 p.
Mosbrugger V. 1992. The evolution of biodiversity - a review of pheho-mena and problems // Rev. Paleobiol. Vol. 1. № 2. P. 533-543.
Newell N. D. 1959. Adequacy of the fossil record // J. Paleontol. Vol.33. № 3. P. 488-499.
Newell N. D. 1967. Revolutions in the history of life // Albretton C. C., Jr. (ed.). Uniformity and simplicity. P. 62-92. (Geol. Soc. Amer. Spec. Paper № 89).
Odin G.-S. 1994. Geological time scale (1994) // C. r. Acad. Sci. Paris. Ser. II. T. 318. P. 59-71.
Odin G.-S., Odin Ch. 1990. Echelle numerique des temps geologiques mise a jour 1990 // Geochronique. № 35. P. 12-15, 17-19.
Patterson C., Smith A. B. 1987. Is the periodicity of extinction a taxono-mic artefact ? // Nature. Vol. 330. № 6145. P. 248-251.
Patterson C., Smith A. B. 1989. Periodicity in extinction: the role of sys-temаtics // Ecology. Vol. 70. № 4. P. 802-811.
Pease C. M. 1985. Biases in the duration and diversities of fossil taxa // Paleobiology. Vol. 11. № 3. P. 272-292.
Peters S. E., Foote M. 2001. Biodiversity in the Phanerozoic: a reinterpre-tation // Paleobiology. Vol. 27. № 4. P. 583-601.
Plotnick R. E., McKinney M. L. Ecosystem organization and extinction dynamics // Palaios. 1993. Vol. 8. № 2. P. 202-212.
Quicke D. L. J. 1996. Principles and techniques of temporary taxonomy. London etc.: Blackie Acad. 311 p.
Raup D. M. 1978. Cohors analysis of generic survivorship // Paleobiolo-gy. Vol. 4. № 4. P. 1-15.
Raup D. M. 1979. Size of the Permo-Triassic bottleneck and its evolutio-nary implications // Science. Vol. 206. № 4415. P. 217-218.
Raup D. M. 1979. Biases in the fossil record of species and genera // Bull. Carnegie Museum Natur. History. № 13. P. 85-91.
Raup D. M. 1986. Biological extinction in Earth history // Science. Vol. 231. № 4745. P. 1528-1533.
Raup D. M., Gould S. J., Schopf T. J. M., Simberloff D. S. 1973. Stochas-tic models of phylogeny and the evolution of diversity // J. Geol. Vol. 81. № 5. P. 525-542.
Raup D. M., Sepkoski J. J., Jr. 1984. Periodicity of extinctions in the geo-logical past // Proc. Nat. Acad. Sci. USA. Biol. sci. Vol. 81. № 2. P. 801-805.
Ross A. J., Jarzembowski E. A. 1993. Arthropoda (Hexapoda; Insecta) // Benton M. J. (ed.). The fossil record (2nd edition). London: Chapman and Hall. P. 363-426.
Schopf T. J. M. 1974. Permo-Triassic extinctions: relation to see-floor spreading // J. Geol. Vol. 82. № 2. P. 129-143.
Schopf T. J. M., Raup D. M., Gould S. J., Simberloff D. S. 1975. Geno-mic versus morphologic rates of evolution: influence of morphologic complexi-ty // Pa-leobiology. Vol. 1. № 1. P. 63-70.
Sepkoski J. J., Jr. 1976. Species diversity in the Phanerozoic: species - area effects // Paleobiology. Vol. 2. № 4. P. 298-303.
Sepkoski J. J., Jr. 1979. A kinetic model of Phanerozoic diversity. II. Early Phanerozoic families and multiple equilibria // Paleobiology. Vol. 5. № 3. P. 222-251.
Sepkoski J. J., Jr. 1982. A compendium of fossil marine animal families // Contrib. in biol. and geol. Milwaukee Public Museum. № 51. 125 p.
Sepkoski J. J., Jr. 1988. Alpha, beta, or gamma: where does all diversity go? // Paleobiology. Vol. 14. № 3. P. 221-234.
Sepkoski J. J., Jr. 1992. A compendium of fossil marine animal families (second edition) // Contrib. in biol. and geol. Milwaukee Public Museum. № 83. 156 p.
Sepkoski J. J., Jr. 1993a. Limits to randomness in paleobiologic models: the case of Phanerozoic species diversity // Acta paleontol. polon. Vol. 38. № 3/4. P. 175-198.
Sepkoski J. J., Jr. 1993b. Ten years in the library: New data confirm pale-ontological patterns // Paleobiology. Vol. 19. № 1. P. 43-51.
Sepkoski J. J., Jr. 1996. Patterns of Phanerozoic extinction: a perspective from global data bases // Walliser O. H. (ed.). Global events and event stratigra-phy in the Phanerozoic: Results of International interdisciplinary cooperation in the IGCP Project 216 "Global biological events in Earth history". Berlin – Hei-delberg: Springer. P. 35-51.
Sepkoski J. J., Jr., Bambach R. K., Raup D. M., Valentine J. W. 1981. Phanerozoic marine diversity and the fossil record // Nature. Vol. 293. № 5832. P. 435-437.
Sepkoski J. J., Jr., Koch C. F. 1996. Evaluating paleontologic data rela-ting to bio-events // Walliser O. H. (ed.). Global events and event stratigraphy in the Phanerozoic: Results of International interdisciplinary cooperation in the IGCP Project 216 "Global biological events in Earth history". Berlin – Heidel-berg: Springer. P. 21-34.
Signor Ph. W. 1985. Real and apparent trends in species richness through time // Valentine J. W. (ed.). Phanerozoic diversity patterns: Profiles in macro-evolution. Princeton: Princeton Univ. Press. P. 129-150.
Signor Ph. W., Lipps J. H. 1982. Sampling bias, gradual extinction pat-terns and catastrophes in the fossil record // Silver L. T., Silver P. H. (eds). Geo-logical implication of impact of large asteroids and comets of the Earth. P. 291-296. (Geol. Soc. Amer. Spec. Paper № 190).
Smith A. B., Patterson C. 1988. The influence of taxonomic method on the perception of patterns of evolution // Evol. Biol. Vol. 23. P. 127-216.
Sneath A., Sokal R. R. 1973. Numerical taxonomy: The principles and practics of numerical classification. San Francisco: Freeman. 573 p.
Sole R. V., Manrubia S., Benton M., Bak P. 1997. Self-similarity of extin-ction statistics in the fossil record // Nature. Vol. 388. № 6644. P. 764-767.
Webb S. D. 1969. Extinction - origination equilibria in Late Cenozoic land mammals of North America // Evolution. Vol. 23. P. 688-702.
Wenz W. 1938-1944. Gastropoda. I. Allgemeiner Teil und Prosobranchia // Handbuch der Palaeozoologie. B. 6. Berlin, 1938. Teil 1. S. 1-240; Teil 2. S. 241-480; 1939. Teil 3. S. 481-720; 1940. Teil 4. S. 721-960; 1941. Teil 5. S. 961-1200; 1943. Teil 6. S. 1201-1506; 1944. Teil 7. S. 1507-1639.
Wenz W., Zilch A. 1960. Gastropoda. II. Euthyneura // Handbuch der Pa-laeozoologie. B. 6. Berlin. Teil 2. 834 S.
Wise K. P., Schopf T. J. M. 1981. Was marine faunal diversity in the Plei-stocene affected by changes in sea level ? // Paleobiology. Vol. 7. № 3. P. 394-399.
<![if !supportFootnotes]>[1]<![endif]> В последнее время попытка изучения зависимости скоростей от разнообразия была сделана А. В. Марковым (2002), однако фактически изучались не скорости, а число по-явившихся и вымерших родов по векам, то же в его предыдущей работе (Марков, 2001), где удельные скорости подменены долями появившихся и вымерших родов.